Daphnia pulex
| Daphnia pulex | |
|---|---|
 | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Subphylum: | Crustacea |
| Class: | Branchiopoda |
| Order: | Cladocera |
| Family: | Daphniidae |
| Genus: | Daphnia |
| Subgenus: | Daphnia |
| Species: | D. pulex |
| Binomial name | |
| Daphnia pulex Leydig, 1860[1][Note 1] | |
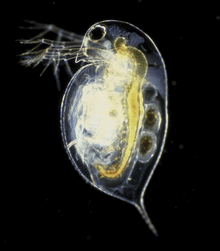
Daphnia pulex (D. pulex) is the most common species of water flea.[3] It has a cosmopolitan distribution: the species is found throughout the Americas, Europe and Australia.[4] It is a model species, and was the first crustacean to have its genome sequenced.
Description
Daphnia pulex is an arthropod whose body segments are difficult to distinguish. They can only be recognised by the appendages they bear (only ever one pair per segment), and by studying the internal anatomy.[5] The head is distinct and is made up of six segments which are fused together even as an embryo. It bears the mouthparts, and two pairs of antennae, the second of which are enlarged into powerful organs used for swimming.[5] There is no clear division between thorax and abdomen, which collectively bear five pairs of appendages.[5] The shell surrounding the animal extends posteriorly into a long or short spine.[6]
Ecology
Daphnia pulex occurs in a wide range of aquatic habitats, although it is most closely associated with small, shaded pools.[7] In oligotrophic lakes, D. pulex has little pigmentation, while it may become bright red in hypereutrophic waters, due to the production of haemoglobin.[7]
Predation
Daphnia are prey for a variety of both vertebrate and invertebrate predators. The role of predation on D. pulex population ecology extensively studied, and has been shown to be a major axis of variation in shaping population dynamics[8] and landscape-level distribution.[9] In addition to the direct population ecological effects of predation, the process contributes to phenotypic evolution in contrasting ways: larger D. pulex are more visible to vertebrate predators, but invertebrate predators are unable to handle larger D. pulex. As a result, larger D. pulex tend to be found with invertebrate predators while smaller size is associated with vertebrate predators.
Similar to some other Daphnia species, the morphology of D. pulex exhibits a plastic response to the presence of predators. Phantom midge larvae (Chaoborus) release kairomones – chemical cues – that induce the development of small jagged protrusions on the head, known as "neck-teeth".[10] Neck-teeth increase survivorship in the presence of the invertebrate predator, but there are costs – longer development time, for example – when those predators are not present.[11]
Ecological stoichiometry
| External identifiers for Daphnia pulex | |
|---|---|
| Encyclopedia of Life | 338871 |
| ITIS | 83874 |
| NCBI | 6669 |
| WoRMS | 148375 |
| Also found in: ADW | |
Daphnia pulex ecology is shaped by nutrient availability and balance, which affects traits that mediate intra- and interspecific interactions. Because nutrients are required for an array of biological processes – for example, amino acid synthesis – the environmental availability of these nutrients regulates downstream organismal characteristics.[12] Low nutrient availability reduces both body size and growth rate, which, as noted above, regulates Daphnia relationships to predators. Daphnia pulex in particular has been an important model species for investigating ecological stoichiometry, demonstrating that pond shading by trees increases nutrient concentrations relative to carbon in algae, which increases D. pulex body size, and therefore competitive ability and susceptibility to predation by vertebrates.[13]
Genomics
Daphnia pulex was the first crustacean to have its genome sequenced.[14][15] Its genome contains 31,000 genes – 8,000 more than are present in the human genome – as a result of extensive gene duplication.[16]
One of the most astonishing features of the D. pulex genome is its compactness: despite being around 200 Mb in size (around 16-fold smaller than the human genome which is 3,200 Mb in size); its 12 chromosomes contain a minimum set of 30,907 predicted protein-coding genes, more than the 20,000–25,000 contained in the human counterpart.[16]
Such an efficient gene packaging is achieved by means of a small intron size. Indeed, whereas the mean protein length in D. pulex is quite similar to that of Drosophila, the average gene size is 1000 bp shorter in D. pulex. As inferred from EST analysis, the average intron size found in D. pulex genes is 170 bp.[16]
On the other hand, the intron density of D. pulex genome is similar to that of Apis mellifera, which in turn is twice of that found in Drosophila.[16]
D. pulex genome has undergone extensive gene duplication followed by rapid paralog divergence and tandem rearrangement. As a result of these processes, around 20% of its gene catalog is composed of tandems consisting of 3–80 paralog genes, many of which are ecoresponsive, that is, they are differentially expressed upon exposure of D. pulex to environmental challenges like biotic or abiotic stress and also fluctuations in light/oxygen levels.[16]
Notes
References
- ↑ Gregorio Fernandez-Leborans; Maria Luisa Tato-Porto (2000). "A review of the species of protozoan epibionts on crustaceans. II. Suctorian ciliates". Crustaceana. 73 (10): 1205–1237. doi:10.1163/156854000505209. JSTOR 20106394.
- ↑ "Daphnia pulex Leydig, 1860". Integrated Taxonomic Information System. Retrieved August 27, 2010.
- ↑ Carrie Miller. "Daphnia pulex". Animal Diversity Web. University of Michigan.
- ↑ "Daphnia pulex". An Image-Based Key To The Zooplankton Of The Northeast (USA). University of New Hampshire.
- 1 2 3 Alexander Ivanovitch Petrunkevitch (1916). "Daphnia pulex". Morphology of Invertebrate Types. pp. 113–121. ISBN 978-0-554-71763-0.
- ↑ Clarence Luther Herrick (2009). "Section 6". A Final Report on the Crustacea of Minnesota. General Books LLC. pp. 21–66. ISBN 978-1-150-02333-0.
- 1 2 "Daphnia pulex". An Image-Based Key To The Zooplankton Of The Northeast (USA). Version 4.0. University of New Hampshire. Retrieved May 12, 2011.
- ↑ Barbara Leoni; Letizia Garibaldi (2009). "Population dynamics of Chaoborus flavicans and Daphnia spp.: effects on a zooplankton community in a volcanic eutrophic lake with naturally high metal concentrations (L. Monticchio Grande, Southern Italy)" (PDF). Journal of Limnology. 68 (1): 37–45.
- ↑ J. H Pantel; T. E Juenger; M. A. Leibold (2011). "Environmental gradients structure Daphnia pulex × pulicaria clonal distribution". Journal of Evolutionary Biology. 24 (4): 723–732. doi:10.1111/j.1420-9101.2010.02196.x.
- ↑ Winfried Lampert; Ulrich Sommer (2007). "Predation". Limnoecology: The Ecology of Lakes and Streams (2nd ed.). Oxford University Press. pp. 162–179. ISBN 978-0-19-921393-1.
- ↑ R. Tollrian (1993). "Neckteeth formation in Daphnia pulex as an example of continuous phenotypic plasticity: morphological effects of Chaoborus kairomone concentration and their quantification". Journal of Plankton Research. 15 (11): 1309–1318. doi:10.1093/plankt/15.11.1309.
- ↑ Robert Warner Sterner; James J. Elser (2002). Ecological Stoichiometry: the Biology of Elements from Molecules to the Biosphere. Princeton University Press. ISBN 978-0-691-07491-7.
- ↑ Spencer R. Hall; Mathew A. Leibold; David A. Lytle; Val H. Smith (2004). "Stoichiometry and planktonic grazer composition over gradients of light, nutrients, and predation risk". Ecology. 85 (8): 2291–2301. doi:10.1890/03-0471.
- ↑ "Daphnia pulex v1.0". DOE Joint Genome Institute. Retrieved 2009-11-29.
- ↑ Florian Odronitz; Sebastian Becker; Martin Kollmar (2009). "Reconstructing the phylogeny of 21 completely sequenced arthropod species based on their motor proteins". BMC Genomics. 10: 173. doi:10.1186/1471-2164-10-173. PMC 2674883
 . PMID 19383156.
. PMID 19383156. - 1 2 3 4 5 John K. Colbourne; Michael E. Pfrender; Donald Gilbert; et al. (2011). "The ecoresponsive genome of Daphnia pulex". Science. 331 (6017): 555–561. Bibcode:2011Sci...331..555C. doi:10.1126/science.1197761. PMC 3529199. PMID 21292972.