Karrikin
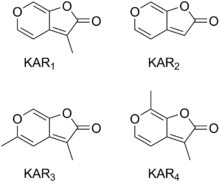
Karrikins are a group of plant growth regulators found in the smoke of burning plant material.[1][2] For many years smoke from wildfires or bushfires was known to stimulate the germination of seeds.[3] In 2004, butenolide was shown to be responsible for this effect.[4] Later, several closely related compounds were discovered in smoke, and are collectively known as karrikins.[2] Six karrikins have been discovered in smoke and are designated KAR1, KAR2, KAR3, KAR4, KAR5 and KAR6, but KAR1 to KAR4 are the most active.[5] The butenolide part of the compound is a 5-membered lactone ring while the other part of the karrikin compound is a 6-membered pyran ring.
Origin
Karrikins are formed by the heating or combustion of carbohydrates including sugars and cellulose.[6] When plant material burns, these carbohydrates convert to karrikins. Plant cell walls are made of polysaccharides including cellulose, and a convenient way to prepare karrikins is to burn paper or dried grass. The pyran part of karrikins is probably directly derived from a pyranose sugar. There is no evidence that karrikins occur naturally in plants, but it has been postulated that karrikin-like molecules do.[7]
Etymology
One of the first recorded Noongar words for 'smoke' from the Perth area in the 1830s, is 'karrik'. This led to these compounds being named karrikins [2][8][9] .
The response to karrikins
Karrikins produced by bushfires occur largely in the ash at the site of the fire. Rains occurring after the fire wash the karrikins into the soil where dormant seeds reside. The karrikins and water can provide a 'wake-up call’ for such seeds, triggering germination of the soil seed bank. The plants that emerge grow quickly, flower and produce new seeds, which fall to the ground. These seeds can remain in the soil for decades, until the next fire produces fresh karrikins. Plants with this lifestyle are known as fire ephemerals. They thrive because the fire removes competing vegetation and provides nutrients and light for the emerging seedlings. Plants in many families respond to smoke and karrikins, suggesting that this response has evolved independently in different groups.[5]
Perception of karrikins
The mode of action of karrikins has been largely determined using the genetic resources of Arabidopsis thaliana. Perception of karrikins by Arabidopsis requires an alpha/beta-fold hydrolase named KARRIKIN-INSENSITIVE-2 (KAI2).[10] The KAI2 protein has a catalytic triad of amino acids which is essential for activity, consistent with the hypothesis that KAI2 hydrolyses its ligand.[11][12] This model is consistent with the perception of the chemically related strigolactone hormones which involves hydrolysis by their receptor protein DWARF14, an alpha/beta hydrolase related to KAI2.[10][13] The question of whether karrikins act directly in plants is controversial. While some studies suggest that karrikins can bind directly to KAI2 protein,[14] others do not support this.[12] It is possible that karrikins produced by wildfires are converted to a different compound by the plant, before interaction with KAI2. The ability of different plants to carry out this conversion could partly explain differences in their ability to respond to karrikins and to smoke.
Karrikin signalling
The activity of karrikins requires an F-box protein named MORE AXILLARY GROWTH-2 (MAX2) in Arabidopsis.[15] This protein is also required for strigolactone signaling in Arabidopsis. Homologs of MAX2 are also required for strigolactone signaling in rice (known as DWARF3) petunia (DAD2) and pea (RMS4). Karrikin signaling also requires a protein named SUPPRESSOR OF MORE AXILARY GROWTH2-1 (SMAX1) [16] which is a homolog of the DWARF53 protein required for strigolactone signaling in rice.[17][18] SMAX1 and DWARF53 proteins could be involved in the control of cellular functions such as transport or transcription.[13] The present model for karrikin and strigolactone signaling involves interaction of KAI2 or DWARF14 with SMAX1 or DWARF53 proteins respectively, which targets those proteins for ubiquitination and destruction.[19]
Effects of karrikins on plant growth
Karrikins not only stimulate seed germination, but are reported to increase seedling vigour.[20] In Arabidopsis karrikins influence seedling photomorphogenesis, resulting in shorter hypocotyls and larger cotyledons. Such responses could provide seedlings with an advantage as they emerge into the post-fire landscape. The KAI2 protein is also required for leaf development, implying that karrikins could influence other aspects of plant growth.
Evolution of karrikin response
The gene for KAI2 protein is present in lower plants including algae and mosses, whereas the DWARF14 protein evolved with seed plants, probably as a result of duplication of KAI2 followed by functional specialisation. Karrikin signaling could have evolved with seed plants as a result of the divergence of KAI2 and DWARF14 functions, possibly during the Cretaceous period when fires were common on Earth.[21]
The endogenous signal for KAI2
Karrikins are produced by wildfires but all seed plants contain KAI2 proteins, raising the question of the usual function of this protein. There is compelling evidence that plants contain an endogenous compound that is perceived by KAI2 to control seed germination and plant development, but this compound is neither a karrikin nor a strigolactone.[12]
References
- ↑ Flematti, Gavin R.; Dixon, Kingsley W; Smith, Steven M. (2015-12-21). "What are karrikins and how were they 'discovered' by plants?". BMC Biology. 13 (1). doi:10.1186/s12915-015-0219-0. PMC 4687367
 . PMID 26689715.
. PMID 26689715. - 1 2 3 Chiwocha, Sheila D. S.; Dixon, Kingsley W.; Flematti, Gavin R.; Ghisalberti, Emilio L.; Merritt, David J.; Nelson, David C.; Riseborough, Julie-Anne M.; Smith, Steven M.; Stevens, Jason C. (2009-10-01). "Karrikins: A new family of plant growth regulators in smoke". Plant Science. 177 (4): 252–256. doi:10.1016/j.plantsci.2009.06.007.
- ↑ HALFORD, BETHANY (2010-04-12). "SMOKE SIGNALS". Chemical & Engineering News Archive. 88 (15): 37–38. doi:10.1021/cen-v088n015.p037. ISSN 0009-2347.
- ↑ Flematti GR, Ghisalberti EL, Dixon KW, Trengove RD (2004). "A compound from smoke that promotes seed germination". Science. 305 (5686): 977. doi:10.1126/science.1099944. PMID 15247439.
- 1 2 Nelson, David C.; Flematti, Gavin R.; Ghisalberti, Emilio L.; Dixon, Kingsley W.; Smith, Steven M. (2012-01-01). "Regulation of Seed Germination and Seedling Growth by Chemical Signals from Burning Vegetation". Annual Review of Plant Biology. 63 (1): 107–130. doi:10.1146/annurev-arplant-042811-105545. PMID 22404467.
- ↑ Flematti, Gavin R.; Scaffidi, Adrian; Dixon, Kingsley W.; Smith, Steven M.; Ghisalberti, Emilio L. (2011-02-23). "Production of the Seed Germination Stimulant Karrikinolide from Combustion of Simple Carbohydrates". Journal of Agricultural and Food Chemistry. 59 (4): 1195–1198. doi:10.1021/jf1041728. ISSN 0021-8561.
- ↑ Gutjahr C et al. (2015). "Rice perception of symbiotic arbuscular mycorrhizal fungi requires the karrikin receptor complex". Science. 350 (6267): 1521–4. doi:10.1126/science.aac9715. PMID 26680197.
- ↑ Flematti, Gavin R.; Ghisalberti, Emilio L.; Dixon, Kingsley W.; Trengove, Robert D. (2009-10-28). "Identification of Alkyl Substituted 2H-Furo[2,3-c]pyran-2-ones as Germination Stimulants Present in Smoke". Journal of Agricultural and Food Chemistry. 57 (20): 9475–9480. doi:10.1021/jf9028128. ISSN 0021-8561.
- ↑ Nelson, David C.; Riseborough, Julie-Anne; Flematti, Gavin R.; Stevens, Jason; Ghisalberti, Emilio L.; Dixon, Kingsley W.; Smith, Steven M. (2009-02-01). "Karrikins Discovered in Smoke Trigger Arabidopsis Seed Germination by a Mechanism Requiring Gibberellic Acid Synthesis and Light". Plant Physiology. 149 (2): 863–873. doi:10.1104/pp.108.131516. ISSN 1532-2548. PMC 2633839. PMID 19074625.
- 1 2 Waters, Mark T.; Nelson, David C.; Scaffidi, Adrian; Flematti, Gavin R.; Sun, Yueming K.; Dixon, Kingsley W.; Smith, Steven M. (2012-04-01). "Specialisation within the DWARF14 protein family confers distinct responses to karrikins and strigolactones in Arabidopsis". Development. 139 (7): 1285–1295. doi:10.1242/dev.074567. ISSN 0950-1991. PMID 22357928.
- ↑ Waters, Mark T.; Scaffidi, Adrian; Flematti, Gavin; Smith, Steven M. (2015-04-05). "Substrate-Induced Degradation of the α/β-Fold Hydrolase KARRIKIN INSENSITIVE2 Requires a Functional Catalytic Triad but Is Independent of MAX2". Molecular Plant. 8 (5): 814–817. doi:10.1016/j.molp.2014.12.020. ISSN 1674-2052. PMID 25698586.
- 1 2 3 Waters, Mark T.; Scaffidi, Adrian; Moulin, Solène L. Y.; Sun, Yueming K.; Flematti, Gavin R.; Smith, Steven M. (2015-07-01). "A Selaginella moellendorffii Ortholog of KARRIKIN INSENSITIVE2 Functions in Arabidopsis Development but Cannot Mediate Responses to Karrikins or Strigolactones". The Plant Cell. 27 (7): 1925–1944. doi:10.1105/tpc.15.00146. ISSN 1532-298X. PMC 4531350. PMID 26175507.
- 1 2 Smith, Steven M; Li, Jiayang (2014-10-01). "Signalling and responses to strigolactones and karrikins". Current Opinion in Plant Biology. SI: Cell signalling and gene regulation. 21: 23–29. doi:10.1016/j.pbi.2014.06.003.
- ↑ Guo, Yongxia; Zheng, Zuyu; Clair, James J. La; Chory, Joanne; Noel, Joseph P. (2013-05-14). "Smoke-derived karrikin perception by the α/β-hydrolase KAI2 from Arabidopsis". Proceedings of the National Academy of Sciences. 110 (20): 8284–8289. doi:10.1073/pnas.1306265110. ISSN 0027-8424. PMC 3657771. PMID 23613584.
- ↑ Nelson, David C.; Scaffidi, Adrian; Dun, Elizabeth A.; Waters, Mark T.; Flematti, Gavin R.; Dixon, Kingsley W.; Beveridge, Christine A.; Ghisalberti, Emilio L.; Smith, Steven M. (2011-05-24). "F-box protein MAX2 has dual roles in karrikin and strigolactone signaling in Arabidopsis thaliana". Proceedings of the National Academy of Sciences. 108 (21): 8897–8902. doi:10.1073/pnas.1100987108. ISSN 0027-8424. PMC 3102411. PMID 21555559.
- ↑ Stanga, John P.; Smith, Steven M.; Briggs, Winslow R.; Nelson, David C. (2013-09-01). "SUPPRESSOR OF MORE AXILLARY GROWTH2 1 Controls Seed Germination and Seedling Development in Arabidopsis". Plant Physiology. 163 (1): 318–330. doi:10.1104/pp.113.221259. ISSN 1532-2548. PMC 3762653. PMID 23893171.
- ↑ Jiang, Liang; Liu, Xue; Xiong, Guosheng; Liu, Huihui; Chen, Fulu; Wang, Lei; Meng, Xiangbing; Liu, Guifu; Yu, Hong (2013-01-01). "DWARF 53 acts as a repressor of strigolactone signalling in rice". Nature. 504 (7480). doi:10.1038/nature12870.
- ↑ Zhou, Feng; Lin, Qibing; Zhu, Lihong; Ren, Yulong; Zhou, Kunneng; Shabek, Nitzan; Wu, Fuqing; Mao, Haibin; Dong, Wei (2013-01-01). "D14–SCFD3-dependent degradation of D53 regulates strigolactone signalling". Nature. 504 (7480). doi:10.1038/nature12878. PMC 4096652. PMID 24336215.
- ↑ Bennett, Tom; Leyser, Ottoline (2014-12-01). "Strigolactone signalling: standing on the shoulders of DWARFs". Current Opinion in Plant Biology. SI: Cell biology. 22: 7–13. doi:10.1016/j.pbi.2014.08.001.
- ↑ van Staden, Johannes; Sparg, Shane G.; Kulkarni, Manoj G.; Light, Marnie E. (2006-08-01). "Post-germination effects of the smoke-derived compound 3-methyl-2H-furo[2,3-c]pyran-2-one, and its potential as a preconditioning agent". Field Crops Research. 98 (2–3): 98–105. doi:10.1016/j.fcr.2005.12.007.
- ↑ He, Tianhua; Pausas, Juli G.; Belcher, Claire M.; Schwilk, Dylan W.; Lamont, Byron B. (2012-05-01). "Fire-adapted traits of Pinus arose in the fiery Cretaceous". New Phytologist. 194 (3): 751–759. doi:10.1111/j.1469-8137.2012.04079.x. ISSN 1469-8137.