Phengaris rebeli
| Phengaris rebeli | |
|---|---|
.jpeg) | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Lepidoptera |
| Family: | Lycaenidae |
| Genus: | Phengaris |
| Species: | P. rebeli |
| Binomial name | |
| Phengaris rebeli (Hirschke, 1904) | |
| Synonyms | |
| |
The Phengaris rebeli (formerly Maculinea rebeli), common name mountain Alcon blue,[1] is a species of butterfly in the Lycaenidae family. It was first found and described in Styria, Austria, on Mount Hochschwab around 1700.[2] Although it was initially classified as a subspecies of P. alcon, a European researcher, Lucien A. Berger, designated it as a separate species in 1946.
Although P. rebeli is found across the Palearctic (see subspecies), it is difficult to determine exactly where they inhabit because of their similarity with P. alcon.[3] In Western Europe they are thought to be concentrated in the Alps and in Southern Europe.
Behavioral ecologists have found its role as a brood parasite to be of particular interest as, unlike many brood parasites, it does not directly oviposit in the hosts' nests. The P. rebeli parasitizes the colony ant species Myrmica schencki as a larva by using chemical mimicry to trick the ants into believing that they are ant larvae; thus, the ants bring the P. rebeli larvae back to their own nests and feed them.[4] It also parasitizes the food plant Gentiana cruciata early in its life cycle,[5] but in turn, is parasitized by the Ichneumon eumerus when it is in the M. schencki's nest.[6] It was placed on the IUCN Red List in 2000 and is classified as a species 'vulnerable' to extinction.[7] Although a separate species from the P. alcon, its DNA genome and appearance are extremely similar, leading many researchers to argue that any differences between the two species are due to intraspecific variation.[2]
Taxonomy
The Phengaris genus is a subsection within the Maculinea genus (explaining why the P. rebeli was formerly known as the M. rebeli) and the Maculinea-Phengaris clade is thought to be a section within Glaucopsyche. There are three groups within this Maculinea-Phengaris clade: the alcon group, the teleius group, and the Arion-Phengaris group.[8]
The groups are divided based on their alternative strategies of parasitizing its host colony (Myrmica ants).[8] The predatory (in which the caterpillar preys on the host ants) and 'cuckoo' (in which the caterpillars feed off of the ants' regurgitation) strategies are derived characteristics from the alcon group, with the predatory strategy having evolved from the teleius group and the cuckoo strategy having evolved from the arion-Phengaris group. Another way these groups are categorized is by whether or not they release a certain chemical (dorsal nectary organ secretions) to attract the attention of the host ant. This trait is not as prominent in ancestors of the alcon group and in the M. nausithous, which is part of the teleius group.[8]
The P. rebeli is most closely related to both species of the Maculinea alcon (the M. alcon alcon and the M. alcon kondakovi) with the M. alcon alcon being more closely related to the P. rebeli than to the M. alcon kondakovi.[8]
Subspecies
- Phengaris rebeli rebeli (Hirschke, 1904) Central and Southern Europe
- Phengaris rebeli cordidula (Jachontov, [1909]) Caucasus Major
- Phengaris rebeli imitator Tuzov, 2000 northern Tian-Shan, Dzhungarsky, Alatau
- Phengaris rebeli kondakovi Kurentzov, 1970 Transbaikalia, Amur Oblast, Ussuri northeast China
- Phengaris rebeli monticola (Staudinger, 1901) Armenia
- Phengaris rebeli ssp. Altai mountains
Appearance
The P. rebeli are noted for their large blue wings, with the males displaying a violet blue shade and the females displaying blue basal areas interspersed with brown spots, similar to other butterflies in its genus.[9] The undersides of the butterfly are a dark brown with small black spots that are circled in white. It can be distinguished from its close relative, the P. alcon, by the broader black margins of the upperside male and the extensive blue basal areas in the females. It has a wingspan of approximately 32–36 mm.[10]
Habitat
It resides in fairly dry areas at lower levels and damp meadows among trees at alpine levels and is found in altitudes of 1000–2000 meters.[9] Although it is found around the globe, populations of P. rebeli are concentrated in the northern part of Portugal and Greece, the western Pyrenees, France, and eastern part of Germany.[9]
Brood parasitism
Discovery of brood parasitism
The Phengaris rebeli is a brood parasite, an organism that manipulates another organism (the host) to raise its offspring; in this case, the P. rebeli parasitizes a particular species of ant, the Myrmica schencki. The P. rebeli was first discovered to be a brood parasite when a researcher observed M. schencki ants bringing the P. rebeli larvae back to their own nest.[4] One of the proposed hypotheses for this parasitism was that P. rebeli larvae released chemicals to confuse the ants into believing they were ant larvae.[4]
It was determined that P. rebeli larvae use chemical mimicry to persuade the M. schencki ants that the P. rebeli are part of the ant brood.[4] Furthermore, M. schencki ants cannot distinguish the physical differences between the P. rebeli larvae and other non-kin brood because the P. rebeli larvae are far more similar to the M. schencki's larvae than to any other ant species larvae.[4]
P. rebeli live in different habitats; therefore, they do not parasitize the same Myrmica ant species.[11] Through observation and experimentation, researchers found that if P. rebeli try to parasitize a different Myrmica ant species than the one they normally do, the Myrmica ants will identify the P. rebeli larvae as intruders and will kill 100% of the P. rebeli larvae. The differences between the P. rebeli are that they synthesize different hydrocarbons, which allow them to mimic different species of Myrmica ants. This explains why they have no success in being mistaken for another species of Myrmica ant and the ensuing 100% mortality rate when the other Myrmica ant species is not fooled.[11]
Acoustics and social rank in host hierarchy
Once the P. rebeli larvae infiltrate the host’s brood, they ascend to the highest social ranks of the host's hierarchy by using acoustics to achieve social acceptance from worker M. schencki ants. P. rebeli larvae and pupa accomplish this by mimicking the sound that the queen of the ant colony makes, both while as a larva and as a pupa in the colony.[12]
While the Myrmica ant colony can identify each other through chemical signaling, social ranks are partially determined by sound acoustics.[13] Therefore, once the P. rebeli begin to mimic the sound of the queen ant, the worker ants begin to treat the P. rebeli as if it were the queen ant.[12] On the other hand, the queen ant treats the P. rebeli larvae and pupae as if they were rivals, as she is the only one in the colony that recognizes that the P. rebeli larvae is not ant larvae.[14]
The most common functions of the queen ant sounds are to recruit workers, smell nestmates, and facilitate oral exchanges of food and pheromones.[14] Most importantly, however, is the fact that distress noises made by the queen causes workers to raise their guard and bolster her protection.
Researchers speculate that acoustical mimicry is related to the level of interaction between the host and parasite. In the Phengaris genus, there are two different strategies: the cuckoo strategy used by P. rebeli and the predatory strategy used by Phengaris arion.[15] In P. rebeli, the Phengaris larvae become integrated into the colony and are attended by worker ants. However, larvae in predatory species prey on the ants’ brood and consequently spend much of their life hiding in pockets of the brood nest.[15]
Integration into host's life
There are two phases for P. rebeli's complete integration into the Myrmica schencki ant colony: initial integration and full integration. In both stages of integration, the P. rebeli caterpillar is brought into the brood nest; however, in full integration, the P. rebeli also achieves its high social status within the host society. That status is crucial for surviving periods of host colony stress such as food shortage.[16]
Studies have shown that the P. rebeli caterpillars benefit most when they parasitize off of the Myrmica schencki ant colony than any other ant colony. When Myrmica ant colonies encounter a period of food shortage, more P. rebeli caterpillars survive than if this food shortage were to occur in any other species of ant colony.[17] This is because the P. rebeli caterpillars have a lower social rank in other ant species compared to their social rank in the M. schencki ant colony.[17] This phenomenon is seen during times of stress, when some of the hungry P. reblei caterpillars secrete compounds to attract attention from the ant colony it has parasitized.[17] While the M. schencki ants are still fooled into believing that the P. rebeli are of their own brood, these compounds do not mimic those of non-host species' societies in other ant species, and thus, results in the P. rebeli being identified as intruders and killed.[17]
On the other hand, the M. schencki prefers to feed the P. rebeli during times of food shortage.[18] Thus, in periods of starvation, P. rebeli caterpillars overall exhibit a higher survival rate than those of the M. schencki larvae.[16]
Polymorphism of growth in larvae
Polymorphism in Lepidoptera is a common occurrence and the P. rebeli caterpillar is no exception as it displays polymorphism in its larval form. The P. rebeli caterpillars have two strategies for living and growing underground: to exist as fast-developing larvae (FDL) or slow-developing larvae (SDL). After the ant brood adopts the FDL, which comprise approximately 25% of the total P. rebeli larvae,[19] the FDL complete growth the following spring and eclose (emerge as an adult from the pupa) in early summer to complete their life cycle.[20] The SDL, which comprise 75% of P. rebeli larvae, do not grow much during the first year, but grow rapidly during the early part of the second summer and remain a second winter within the ant colonies.[20] While both larva types ultimately form similar-sized pupae, their polymorphic growth rates could indicate alternative fitness strategies and different ways to exploit the M. schencki's food resources.[20]
Other researchers hypothesize that another alternative growth strategy will evolve in the P. rebeli, in which the P. rebeli will parasitize the M. schencki ants for an even longer period of time than the slow developing larvae. Most researchers, however, find this hypothesis highly unlikely because it is not an evolutionarily stable strategy. At the rate the P. rebeli parasitizes the M. schencki ants, the host colony is unlikely to last as a viable food source for more than two years (the average lifespan of the longer polymorph of P. rebeli). Once the host colony reaches its lifespan of two years and dies, the P. rebeli loses its food source and dies as well.[19]
The P. rebeli's developmental rate shows great phenotypic plasticity (changing its developmental rate in response to changes in the environment), as it develops very quickly in the lab and in the Pyrenees and Southern Alps.[21] Scientists hypothesize that this plasticity is due to warm conditions and more light exposure, which affects larval development. Abundant food resources are also thought to play a part in quickened development.[21]
Studies have shown that smaller-sized P. rebeli from Hungary normally eclose at the end of June or mid-July; however, when these populations are studied in laboratories, they do not eclose until late August.[21] This has led researchers to speculate that P. rebeli larvae that do not receive adequate food can still fully develop in one year and act as a functioning adult. However, they are unlikely to be bivoltine (producing two broods in one season). This hypothesis is supported by the fact that certain researchers claim that P. rebeli larvae can have continuous development.[21]
Vestigial mutualism
P. rebeli pupae prey upon their broodmates, while producing sugar-rich secretions which worker ants imbibe. In one experiment, P. rebeli specimens which consumed ant larvae developed more quickly than those who did not. However, despite their nourishing offering to the community, they invariably imposed a net loss in the survival rates of workers and broodmates, demonstrating that the species is parasitic (rather than mutualistic) at all stages in its host colony.[22]
Relationship with host plant Gentiana cruciata
Besides parasitizing the host ant, P. rebeli also parasitizes the grassland plant Gentiana cruciata as a source of nutrition and feeds off of its flowers and developing fruits.[5]
The larger the population size of the G. cruciata, the more flowers and seeds the plant produces; however, there is also a positive link between the population size of the G. cruciata and the frequency of P. rebeli parasitizing the plant.[5] Researchers are particularly interested in the G. cruciata plant because both the P. rebeli and G. cruciata are endangered.[5] As a result, lepidopterists believe that conserving the P. rebeli requires focusing conservation efforts on preserving G. cruciata as well.[5]
Researchers discovered that where the P. rebeli lay their eggs is based on the size of the G. cruciata leaves and not upon the location of the closest Myrmica ant colony.[23] Evidence for P. rebeli's oviposition depending upon the range of the food plant rather than the range of the ants is supported by the timing of its oviposition. P. rebeli lays their eggs during the warm summer season, when Myrmica ants are most likely to be underground.[23] Researchers also observed females did not base their oviposition on where they found Myrmica nests.[23]
Relationship with the parasitic wasp, Ichneumon eumerus
Though P. rebeli parasitizes the Myrmica schencki ants and Gentiana cruciata food plant, it also acts as a host to the parasitoid ichneumon wasp (Ichneumon eumerus).[6] The I. eumerus is a wasp that attempts to parasitize the P. rebeli after it has been introduced to the Myrmica ants' nests. The adult wasp searches for Myrmica ant nests by responding strongly to the scent of Myrmica schencki ants, the species that P. rebeli most commonly and successfully parasitizes.[6]
After locating the M. schnecki nest, the wasp enters only the nests that contain P. rebeli caterpillars.[6] It is able to enter the nest without being swarmed because it releases a chemical that causes the worker ants to attack one another instead of concentrating all their efforts on the wasp. Once the wasp reaches the caterpillars, it marks the individuals most likely to survive by ovipositing in them. Once the wasp's eggs hatch from the caterpillar's body, the newborn offspring feast on the caterpillar's carcass.[6]
Conflicts over classification
Several researchers argue that the P. rebeli has not evolved into a separate species from the Phengaris alcon. Researchers have determined that both species share a similar adult morphology, DNA and allozymes. They found that any variation between the two species is due to intraspecific variation, meaning that they are not distinct species.[2] Furthermore, they both have similar methods of parasitizing the host ant: they act as parasitic "cuckoos" within the ant nest and feed upon the ants' regurgitation.[24] This is compared to the alternative method of predation that other butterflies, such as the Maculinea arion utilize to parasitize the host ant.[24]
To test this hypothesis, researchers examined how each species utilized the host ants.[25] The M. schencki, M. sabuleti are parasitized by P. rebeli while the M. salina and M. vandeli are parasitized by the P. alcon. The M. scabrinodis is parasitized by both the P. rebeli and the P. alcon; however, the P. alcon is far more likely to parasitize M. scabrinodis than the P. rebeli.[25] Researchers found that in general, P. alcon was far more likely to parasitize the host ant than P. rebeli; however, ant nests that supported the P. alcon were much smaller and supported lower populations than ant nests that supported P. rebeli.[25]
Conservation status
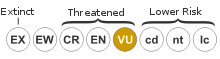
The P. rebeli has been rigorously studied in Europe because it has priority conservation status and was classified as "vulnerable" in 2000 by the IUCN Red List of Threatened Species, which is described to be "the world's most comprehensive inventory" that lists the conservation status of plants and animals.[7] It was first brought to the IUCN's attention and listed on the IUCN Red List of Threatened Species in 1986. The reason for being categorized as "vulnerable" is that despite its wide distribution, it has decreased by 20-50% over the last 25 years and has reached extinction in at least one country.
The reasons for this drastic population reduction are thought to be due to agricultural improvements, abandonment of extensive management, and deforestation.[7] As the area of the Gentiana cruciata, the food plant that the P. rebeli depends on, decreases, the P. rebeli population also decreases.[7]
Gallery
 Eggs on host plant
Eggs on host plant Pupa in ant nest
Pupa in ant nest
References
- ↑ "Maculinea rebeli". UniProt Consortium. Retrieved 2009-02-09.
- 1 2 3 Steiner, F; , Schlick-Steiner B.C. , Hçttinger H. , Nikiforov A. , Moder K. and Christian E. (2006). "Maculinea alcon and M. rebeli (Insecta: Lepidoptera: Lycaenidae) – one or two alcon blues? Larval cuticular compounds and egg morphology of East Austrian populations". Ann. Naturhist. Mus. Wien. 107B: 165–180. JSTOR 41767345.
- ↑ Steiner, F.M; B.C . Schlick-Steiner , H . Höttinger 1 , A . Nikiforov , K . Moder & E . Christian (2006). "Geographical versus food plant differentiation in populations of Maculinea alcon (Lepidoptera: Lycaenidae) in Northern Hungary" (PDF). Ann. Naturhist. Mus. Wien. 107B: 165–180. Retrieved 23 October 2013.
- 1 2 3 4 5 Akino, T; JJ Knapp; JA Thomas; GW Elmes (1999). "Chemical mimicry and host specificity in the butterfly Maculinea rebeli, a social parasite of Myrmica ant colonies". Proceedings of the Royal Society B. 266 (1427): 1419–1426. doi:10.1098/rspb.1999.0796. Retrieved 28 September 2013.
- 1 2 3 4 5 Kéry, M; Matthies, D & Fischer, M (2001). "The effect of plant population size on the interactions between the rare plant Gentiana cruciata and its specialized herbivore Maculinea rebeli". Journal of Ecology. 89 (3): 418–427. doi:10.1046/j.1365-2745.2001.00550.x. Retrieved 19 October 2013.
- 1 2 3 4 5 Hochberg, M; Elmes, G. W.; Thomas, J. A.; Clarke, R. T (1996). "Mechanisms of local persistence in coupled host-parasitoid associations: the case model of Maculinea rebeli and Ichneumon eumerus". Philosophical Transactions: Biological Sciences. 351 (1348): 1713–1724. Bibcode:1996RSPTB.351.1713H. doi:10.1098/rstb.1996.0153. Retrieved 19 October 2013.
- 1 2 3 4 "Maculinea rebeli". Retrieved 19 October 2013.
- 1 2 3 4 Pech, Pavel; Zdene ˇ k Fric, Martin Konvic ˇ ka and Jan Zrzavy; Konvicka, Martin; Zrzavy, Jan (2004). "Phylogeny of Maculinea blues (Lepidoptera: Lycaenidae) based on morphological and ecological characters: evolution of parasitic myrmecophily" (PDF). Cladistics. 20 (4): 362–375. doi:10.1111/j.1096-0031.2004.00031.x. Retrieved 19 October 2013.
- 1 2 3 Rowlings, Matt. "Maculinea rebeli". Retrieved 19 October 2013.
- ↑ Karaman, Seyfi. "Mountain Alcon Blue". Retrieved 2 November 2013.
- 1 2 Thomas, J; Elmes, G. W., Sielezniew, M., Stankiewicz-Fiedurek, A., Simcox, D. J., Settele, J., & Schönrogge K; Sielezniew, M.; Stankiewicz-Fiedurek, A.; Simcox, D. J.; Settele, J.; Schonrogge, K. (2013). "Mimetic host shifts in an endangered social parasite of ants" (PDF). Proceedings of the Royal Society B. 280 (1751): 20122336. doi:10.1098/rspb.2012.2336. PMID 23193127. Retrieved 19 October 2013.
- 1 2 Thomas, JA; Schonrogge K; Bonelli S; Barbero F; Balletto E (2010). "Corruption of ant acoustical signals by mimetic social parasites: Maculinea butterflies achieve elevated status in host societies by mimicking the acoustics of queen ants". Communicative & Integrative Biology. 3 (2): 1–4. doi:10.4161/cib.3.2.10603. PMC 2889977
 . PMID 20585513. Retrieved 28 September 2013.
. PMID 20585513. Retrieved 28 September 2013. - ↑ Barbero, Francesca; Thomas JA; Bonelli S; Balletto E; Schonrogge K (2009). "Acoustical mimicry in a predatory social parasite of ants". Journal of Experimental Biology. 212 (Pt 24): 4084–4090. doi:10.1242/jeb.032912. PMID 19946088. Retrieved 28 September 2013.
- 1 2 Barbero, Francesca; Thomas JA; Bonelli S; Balletto E; Schonrogge K (2009). "Queen ants make distinctive sounds that are mimicked by a butterfly social parasite". Science. 373 (5915): 782–785. Bibcode:2009Sci...323..782B. doi:10.1126/science.1163583. Retrieved 28 September 2013.
- 1 2 Barbero, Francesca; Patricelli D; Witek M; Balletto E; Casacci LP; Sala M; Bonelli S (2012). "Myrmica Ants and Their Butterrfly Parasites with Special Focus on the Acoustic Communication". Psyche. 2012 (725237): 1–11. doi:10.1155/2012/725237. Retrieved 28 September 2013.
- 1 2 Elmes, GW; Wardlaw JC; Schonrogge K; Thomas JA; Clarke, R.T. (2003). "Food stress causes differential survival of socially parasitic larvae of Maculinea rebeli (Lepidoptera, Lycaenidae) integrated in colonies of host and non- host Myrmica species (Hymenoptera, Formicidae)". Ent Exp et Appl. 110: 53–63. doi:10.1111/j.0013-8703.2004.00121.x. Retrieved 23 October 2013.
- 1 2 3 4 Schonrogge, K; Wardlaw JC; Peters AJ; Everett S; Thomas JA; Elmes GW (2004). "Changes in chemical signature and host specificity from larval retrieval to full social integration in the myrmecophilous butterfly Maculinea rebeli". Journal of Chemical Ecology. 30 (1): 91–107. doi:10.1023/b:joec.0000013184.18176.a9. PMID 15074659. Retrieved 28 September 2013.
- ↑ Elmes, GW; Thomas JA; Wardlaw JC (1991). "Larvae of Maculinea rebeli, a large blue butterfly, and their Myrmica host ants: wild adoption and behavior in ant-nests". Journal of Zoology. 224 (3): 447–460. doi:10.1111/j.1469-7998.1991.tb04775.x. Retrieved 28 September 2013.
- 1 2 Thomas, JA; Elmes GW; Wardlaw JC (1998). "Polymorphic growth in larvae of the butterfly Maculinea rebeli, a social parasite of Myrmica ant colonies" (PDF). Proceedings of the Royal Society. 265 (1408): 1895–1901. doi:10.1098/rspb.1998.0517. Retrieved 28 September 2013.
- 1 2 3 Hovestadt, T; Mitesser O; Elmes GW; Thomas JA; Hochberg ME (2007). "An Evolutionarily Stable Strategy model for the evolution of dimorphic development in the butterfly Maculinea rebeli, a social parasite of Myrmica ant colonies". The American Naturalist. 169 (4): 466–480. doi:10.1086/512134. JSTOR 512134. PMID 17269113.
- 1 2 3 4 Tartally, Andras (2005). "Accelerated development of Maculinea rebeli larvae under artificial conditions (Lycaenidae)" (PDF). 27 (4): 303–308. Retrieved 19 October 2013.
- ↑ Wardlaw, JC; Thomas JA; Elmes GW (April 2000). "). "Do Maculinea rebeli caterpillars provide vestigial mutualistic benefits to ants when living as social parasites inside Myrmica ant nests? (Abstract)"". Entomologia Experimentalis et Applicata. 95 (1): 97–103. doi:10.1046/j.1570-7458.2000.00646.x. Retrieved 22 October 2013.
- 1 2 3 Thomas, J; Elmes, G (2001). "Foodplant niche selection rather than the presence of ant nests explains oviposition patterns in the myrmecophilous butterfly genus Maculinea" (PDF). Proceedings of the Royal Society of Lond, Series B. 268 (1466): 471–477. doi:10.1098/rspb.2000.1398. Retrieved 19 October 2013.
- 1 2 Clarke, R; Mouquet, N.; Thomas, J.; Hochberg, M.; Elmes, G.; Tesar, D.; Singer, A.; Hale, J. (2005). "Modelling the local population dynamics of Maculinea and their spatial interactions with their larval foodplant and Myrmica ant species". Retrieved 19 October 2013.
- 1 2 3 Tartally, A; Nash, D. R.; Lengyel, S.; Varga, Z (2008). "Patterns of host ant use by sympatric populations of Maculinea alcon and M.'rebeli'in the Carpathian Basin". Insectes Sociaux. 55 (4): 370–381. doi:10.1007/s00040-008-1015-4. Retrieved 19 October 2013.
External links
| Wikimedia Commons has media related to Maculinea rebeli. |