Triple-resonance nuclear magnetic resonance spectroscopy
Triple resonance experiments are a set of multi-dimensional nuclear magnetic resonance spectroscopy (NMR) experiments that link three types of atomic nuclei, most typically consisting of 1H, 15N and 13C. These experiments are often used to assign specific resonance signals to specific atoms in an isotopically enriched protein. The technique was first described in papers by Ad Bax, Mitsuhiko Ikura and Lewis Kay in 1990,[1][2] and further experiments were then added to the suite of experiments. Many of these experiments have since become the standard set of experiments used for the sequencing assignment of resonances of protein necessary for the determination of protein structure by NMR, and are an integral part of solution NMR study of proteins.[3]
Background
There are two main methods of determining protein structure on the atomic level. The first of these is by X-ray crystallography, starting in 1958 when the crystal structure of myoglobin was determined. The second method is by NMR, which began in the 1980s when Kurt Wüthrich outlined the framework for NMR structure determination of proteins, and solved the solution structure of small globular proteins.[4] Structural determination of protein by NMR however requires the assignment of the NMR signals from the many nuclei in the protein. The early method of protein NMR relied on proton-based homonuclear NMR spectroscopy where the size of the protein that may be determined is limited to ~10 KDa. This limitation is due to the presence of greater number of protons in larger protein which results in overcrowding of the resonances, as well as the broadening of signal that comes with increasing size of protein, making resonance assignment difficult. These problems may be alleviated by using heteronuclear NMR spectroscopy which allows the proton spectrum to be edited with respect to the 15N and 13C chemical shifts, as well as increasing the number of dimension of the spectrum. In 1990, Ad Bax and coworkers developed the triple resonance technology and experiments on proteins isotopically-labelled with 15N and 13C,[1] with the result that the spectra are dramatically simplified, greatly facilitating the process of resonance assignment, and increasing the size of the protein that may be determined by NMR.
These triple resonance experiments utilize the relatively large magnetic couplings between certain pairs of nuclei to establish their connectivity. Specifically, the 1JNH, 1JCH, 1JCC, and 1JCN couplings are used to establish the scalar connectivity pathway between nuclei. The magnetization transfer process takes place through multiple, efficient one-bond magnetization transfer steps, rather than a single step through the smaller and variable 3JHH couplings. The relatively large size and good uniformity of the one-bond couplings allowed the design of efficient magnetization transfer schemes that are effectively uniform across a given protein, nearly independent of conformation.[3] Triple resonance experiments involving 31P may also be use for nucleic acid studies.[5]
Suite of experiments
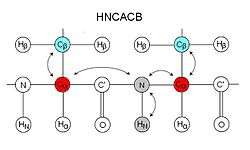

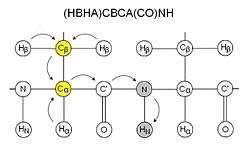
These experiments are typically named by the nuclei involved in the experiment. The nuclei in the name are ordered in the same sequence as in the path of magnetization transfer, those nuclei placed within parentheses are involved in the magnetization transfer pathway but are not recorded. For reason of sensitivity, these experiments generally start on a proton and end on a proton, therefore many of these experiments are what's called "out-and-back" experiments where, although not indicated in the name, the magnetization is transferred back to the starting proton for signal acquisition.
Some of the experiments are used in tandem for the resonance assignment of protein, for example HNCACB, is normally used together with CBCA(CO)NH as a pair of experiments. Not all these experiments need to be recorded for sequential assignment (it can be done with as few as two), however extra pairs of experiment are useful for independent assessment of the correctness of the assignment, and the redundancy of information may be necessary when there is ambiguity in the assignments. Other experiments are also necessary to fully assign the side chain resonances.
TROSY version of some of these experiments exist for improvement in sensitivity.[6]
HNCO
The experiment provides the connectivities between the amide of a residue with the carbonyl carbon of the preceding residues.[2] It is the most sensitive of the triple resonance experiments. The sidechains carboxamides of asparagine and glutamine are also visible in this experiment. Additionally, the guanidino group of arginine, which has similar coupling constant to the carboxamide group, may also appear in this spectrum. This experiment is sometimes used together with HN(CA)CO.
HN(CA)CO
Here, the amide resonance of a residue is correlated with the carbonyl carbon of the same residue, as well as that of the preceding residues. The intra-residue resonances are usually stronger than the inter-residues one.[7]
HN(CO)CA
This experiment correlates the resonances of the amide of a residue with the Cα of the preceding residue. This experiment is often used together with HNCA.[8]
HNCA
This experiment correlates the chemical shift of amide of a residue the Cα of the same residue as well as those of the preceding residue.[2] Each strip gives two peaks, the inter and intra-residue Cα peaks. Peak from the preceding Cα may be identified from the HN(CO)CA experiment which give only the inter-residue Cα.
CBCA(CO)NH
CBCA(CO)NH, or alternatively HN(CO)CACB, correlates the resonances of the amide of a residue with the Cα and Cβ of the preceding residue.[9] Two peaks corresponding to the Cα and Cβ are therefore visible for each residue. This experiment is normally used together with HNCACB. The sidechain carboxamide of glutamines and asparagines also appear in this spectra in this experiment. CBCA(CO)NH is sometimes more precisely called (HBHA)CBCA(CO)NH as it starts with aliphatic protons and ends on an amide proton, and is therefore not an out-and-back experiment like HN(CO)CACB.
HNCACB
HNCACB, or alternatively CBCANH, correlates the chemical shift of amide of a residue the Cα and Cβ of the same residue as well as those of the preceding residue.[10] In each strip, four peaks may be visible – 2 from the same residue and 2 from the preceding residue. Peaks from the preceding residue are usually weaker, and may be identified using CBCA(CO)NH. In this experiment, the Cα and Cβ peaks are in opposite phase, i.e. if Cα appears as a positive peak, then Cβ will be negative, making identification of Cα and Cβ straightforward. The extra information of Cβ from the CBCA(CO)NH/HNCACB set of experiments makes identification of residue type easier than HN(CO)CA/HNCA, however the HNCACB is a less sensitive experiment and may be unsuitable for some proteins.
The CBCANH experiment is less suitable for larger protein as it is more susceptible to the line-width problem than HNCACB.
CBCACO(CA)HA
This experiment provides the connectivities between the Cα and Cβ with the carbonyl carbon and Hα atoms within the same residue.[11] The sidechain carboxyl group of aspartate and glutamate may appear weakly in this spectrum.
CC(CO)NH
This experiment provides connectivities between the amide of a residue and the aliphatic carbon atoms of the preceding residue.[12]
H(CCO)NH
This experiment provides connectivities between the amide of a residue and the hydrogen atoms attached to the aliphatic carbon of the preceding residue.
HBHA(CO)NH
This experiment correlates the amide resonance to the Hα and Hβ of the preceding residue.[13]
Sequential assignment
Pairs of experiments are normally used for sequential assignment, for example, the HNCACB and CBCA(CO)NH pair, or HNCA and HNC(CO)CA. The spectra are normally analyzed as strips of peaks, and strips from the pair of experiments may be presented together side by side or as an overlay of two spectra. In the HNCACB spectra 4 peaks are usually present in each strip, the Cα and Cβ of one residue as well as those of its preceding residue. The peaks from the preceding residue can be identified from the CBCA(CO)NH experiment. Each strip of peaks can therefore be linked to the next strip of peaks from an adjoining residue, allowing the strips to be connected sequentially. The residue type can be identified from the chemical shifts of the peaks, some, such as serine, threonine, glycine and alanine, are however easier to identify then others. The resonances can then be assigned by comparing the sequence of peaks with the amino acid sequence of the protein.
References
- 1 2 Ikura M; Kay LE; Bax A (1990). "A novel approach for sequential assignment of 1H, 13C, and 15N spectra of proteins: heteronuclear triple-resonance three-dimensional NMR spectroscopy. Application to calmodulin". Biochemistry. 29 (19): 4659–67. doi:10.1021/bi00471a022. PMID 2372549.
- 1 2 3 Lewis E Kay; Mitsuhiko Ikura; Rolf Tschudin, Ad Bax (1990). "Three-dimensional triple-resonance NMR spectroscopy of isotopically enriched proteins". Journal of Magnetic Resonance. 89 (3): 496–514. Bibcode:1990JMagR..89..496K. doi:10.1016/0022-2364(90)90333-5.
- 1 2 Ad Bax (2011). "Triple resonance three-dimensional protein NMR: before it became a black box". Journal of Magnetic Resonance. 213 (2): 442–5. Bibcode:2011JMagR.213..442B. doi:10.1016/j.jmr.2011.08.003. PMC 3235243
 . PMID 21885307.
. PMID 21885307. - ↑ Kurt Wüthrich (2001). "The way to NMR structures of proteins". Nature Structural Biology. 8 (11): 923–925. doi:10.1038/nsb1101-923. PMID 11685234.
- ↑ Gabriele Varani; Fareed Aboul-ela; Frederic Allain & Charles C. Gubser (1995). "Novel three-dimensional 1H−13C−31P triple resonance experiments for sequential backbone correlations in nucleic acids". Journal of Biomolecular NMR. 5 (3): 315–320. doi:10.1007/BF00211759.
- ↑ Michael Salzmann; Gerhard Wider; Konstantin Pervushin; Hans Senn & Kurt Wu1thrich (1999). "TROSY-type Triple-Resonance Experiments for Sequential NMR Assignments of Large Proteins" (PDF). Journal of the American Chemical Society. 121 (4): 844–848. doi:10.1021/ja9834226.
- ↑ Robert T Clubb; V Thanabal; Gerhard Wagner (1992). "A constant-time three-dimensional triple-resonance pulse scheme to correlate intraresidue 1HN, 15N, and 13C′ chemical shifts in 15N/13C-labelled proteins". Journal of Magnetic Resonance. 97 (1): 213–217. Bibcode:1992JMagR..97..213C. doi:10.1016/0022-2364(92)90252-3.
- ↑ Ad Bax & Mitsuhiko Ikura (1991). "An efficient 3D NMR technique for correlating the proton and 15N backbone amide resonances with the α-carbon of the preceding residue in uniformly 15N/13C enriched proteins". Journal of Biomolecular NMR. 1 (1): 99–104. doi:10.1007/BF01874573. PMID 1668719.
- ↑ Stephan Grzesiek, Ad Bax (1992). "Correlating backbone amide and side chain resonances in larger proteins by multiple relayed triple resonance NMR". Journal of the American Chemical Society. 114 (16): 6291–6293. doi:10.1021/ja00042a003.
- ↑ Stephan Grzesiek, Ad Bax (1992). "An efficient experiment for sequential backbone assignment of medium-sized isotopically enriched proteins". Journal of the Magnetic Resonance. 99 (1): 201–207. Bibcode:1992JMagR..99..201G. doi:10.1016/0022-2364(92)90169-8.
- ↑ Kay, Lewis E. (1993). "Pulsed-field gradient-enhanced three-dimensional NMR experiment for correlating 13Cα/β, 13C', and 1Hα chemical shifts in uniformly carbon-13-labeled proteins dissolved in water". Journal of the American Chemical Society. 115 (5): 2055–2058. doi:10.1021/ja00058a072.
- ↑ S. Grzesiek; J. Anglister; A. Bax (1993). "Correlation of Backbone Amide and Aliphatic Side-Chain Resonances in 13C/15N-Enriched Proteins by Isotropic Mixing of 13C Magnetization". Journal of the Magnetic Resonance, series B. 101 (1): 114–119. Bibcode:1993JMRB..101..114G. doi:10.1006/jmrb.1993.1019.
- ↑ Stephan Grzesiek & Ad Bax (1993). "Amino acid type determination in the sequential assignment procedure of uniformly 13C/15N-enriched proteins". Journal of Biomolecular NMR. 3 (2): 185–204. doi:10.1007/BF00178261. PMID 8477186.
External links
- Triple resonance experiments for proteins
- Introduction to 3D Triple Resonance Experiments
- Protein NMR – A Practical Guide