Aphid
| Aphids Temporal range: Permian–present | |
|---|---|
-PLoS.jpg) | |
| Pea aphids, Acyrthosiphon pisum Nymph aphids surrounding the mother aphid were produced parthenogenetically and viviparously; sexual reproduction can be induced by shorter amounts of daylight. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hemiptera |
| Suborder: | Sternorrhyncha |
| Superfamily: | Aphidoidea Geoffroy, 1762 |
Aphids, also known as plant lice and in Britain and the Commonwealth as greenflies, blackflies, or whiteflies (not to be confused with "jumping plant lice" or true whiteflies), are small sap-sucking insects, and members of the superfamily Aphidoidea.[1] Many species are green but other commonly occurring species may be white and wooly or black. Aphids are among the most destructive insect pests on cultivated plants in temperate regions.[1] The damage they do to plants has made them enemies of farmers and gardeners around the world. From a zoological standpoint they are a highly successful group of organisms.[2] Their success is due in part to the asexual reproductive capabilities of some species.
About 4,400 species are known, all included in the family Aphididae.[3] Around 250 species are serious pests for agriculture and forestry as well as an annoyance for gardeners. They vary in length from 1 to 10 millimetres (0.04 to 0.39 in).
Natural enemies include predatory ladybugs, hoverfly larvae, parasitic wasps, aphid midge larvae, crab spiders, lacewings, and entomopathogenic fungi such as Lecanicillium lecanii and the Entomophthorales.
Distribution
Aphids are distributed worldwide, but are most common in temperate zones. In contrast to many taxa, aphid species diversity is much lower in the tropics than in the temperate zones. They can migrate great distances, mainly through passive dispersal by riding on winds. For example, the currant-lettuce aphid, Nasonovia ribisnigri, is believed to have spread from New Zealand to Tasmania in this way.[4] Aphids have also been spread by human transportation of infested plant materials.
Taxonomy
Aphids are in the superfamily Aphidoidea in the Sternorrhyncha division of the order Hemiptera. Late 20th-century reclassification within the Hemiptera reduced the old taxon "Homoptera" to two suborders: Sternorrhyncha (e.g., aphids, whiteflies, scales, psyllids, etc.) and Auchenorrhyncha (e.g., cicadas, leafhoppers, treehoppers, planthoppers, etc.) with the suborder Heteroptera containing a large group of insects known as the true bugs. Early 21st-century reclassifications substantially rearranged the families within Aphidoidea: some old families were reduced to subfamily rank (e.g., Eriosomatidae), and many old subfamilies were elevated to family rank. The most recent authoritative classifications place all extant taxa into a single large family Aphididae.[3] Despite their names, taxonomically, the woolly conifer aphids like the pine aphid, the spruce aphid, and the balsam woolly aphid are not true aphids, but adelgids, and lack the cornicles of true aphids.
Relation to phylloxera and adelgids
Aphids, adelgids, and phylloxerids are very closely related, and are all within the suborder Sternorrhyncha, the plant-sucking bugs. They are either placed in the insect superfamily Aphidoidea[5] or into the superfamily Phylloxeroidea which contains the family Adelgidae and the family Phylloxeridae.[6]
Like aphids, phylloxera feed on the roots, leaves, and shoots of grape plants, but unlike aphids, do not produce honeydew or cornicle secretions.[7] Phylloxera (Daktulosphaira vitifoliae) are insects which caused the great French wine blight that devastated European viticulture in the 19th century.
Similarly, adelgids also feed on plant phloem. Adelgids are sometimes described as aphids, but are more properly classified as aphid-like insects, because they have no cauda or cornicles.[1]
Anatomy

Most aphids have soft bodies, which may be green, black, brown, pink, or almost colourless. Aphids have antennae with as many as six segments.[1] They feed themselves through sucking mouthparts called stylets, enclosed in a sheath called a rostrum, which is formed from modifications of the mandible and maxilla of the insect mouthparts.[8] They have long, thin legs and two-jointed, two-clawed tarsi.
Most aphids have a pair of cornicles (or "siphunculi"), abdominal tubes through which they exude droplets of a quick-hardening defensive fluid[8] containing triacylglycerols, called cornicle wax. Other defensive compounds can also be produced by some types of aphids.[1]
Aphids have a tail-like protrustion called a cauda above their rectal apertures. They have two compound eyes, and an ocular tubercle behind and above each eye, made up of three lenses (called triommatidia).[6][9]
When host plant quality becomes poor or conditions become crowded, some aphid species produce winged offspring, "alates", that can disperse to other food sources. The mouthparts or eyes are smaller or missing in some species and forms.[1]
Diet
Many aphid species are monophagous (that is, they feed on only one plant species). Others, like the green peach aphid Myzus persicae, feed on hundreds of plant species across many families.
Aphids passively feed on sap of phloem vessels in plants, as do many of their fellow members of Hemiptera such as scale insects and cicadas. Once a phloem vessel is punctured, the sap, which is under high pressure, is forced into the aphid's food canal. Occasionally, aphids also ingest xylem sap, which is a more dilute diet than phloem sap as the concentrations of sugars and amino acids are 1% of those in the phloem.[10][11] Xylem sap is under negative hydrostatic pressure and requires active sucking, suggesting an important role in aphid physiology.[12] As xylem sap ingestion has been observed following a dehydration period, aphids are thought to consume xylem sap to replenish their water balance; the consumption of the dilute sap of xylem permitting aphids to rehydrate.[13] However, recent data showed aphids consume more xylem sap than expected and they notably do so when they are not dehydrated and when their fecundity decreases.[14] This suggests aphids, and potentially, all the phloem-sap feeding species of the order Hemiptera, consume xylem sap for another reason than replenishing water balance.
Xylem sap consumption may be related to osmoregulation.[14] High osmotic pressure in the stomach, caused by high sucrose concentration, can lead to water transfer from the hemolymph to the stomach, thus resulting in hyperosmotic stress and eventually to the death of the insect. Aphids avoid this fate by osmoregulating through several processes. Sucrose concentration is directly reduced by assimilating sucrose toward metabolism and by synthesizing oligosaccharides from several sucrose molecules, thus reducing the solute concentration and consequently the osmotic pressure.[15][16] Oligasaccharides are then excreted through honeydew, explaining its high sugar concentrations, which can then be used by other animals such as ants. Furthermore, water is transferred from the hindgut, where osmotic pressure has already been reduced, to the stomach to dilute stomach content.[17] Eventually, aphids consume xylem sap to dilute the stomach osmotic pressure.[14] All these processes function synergetically, and enable aphids to feed on high-sucrose-concentration plant sap, as well as to adapt to varying sucrose concentrations.
Plant sap is an unbalanced diet for aphids, as it lacks essential amino acids, which aphids, like all animals, cannot synthesise, and possesses a high osmotic pressure due to its high sucrose concentration.[11][18] Essential amino acids are provided to aphids by bacterial endosymbionts, harboured in special cells, bacteriocytes.[19] These symbionts recycle glutamate, a metabolic waste of their host, into essential amino acids.[20][21]
As they feed, aphids often transmit plant viruses to the plants, such as to potatoes, cereals, sugarbeets, and citrus plants.[8] These viruses can sometimes kill the plants.
Symbioses
Ant mutualism


Some species of ants farm aphids, protecting them on the plants they eat, eating the honeydew the aphids release from the terminations of their alimentary canals. This is a mutualistic relationship. These dairying ants milk the aphids by stroking them with their antennae.[Note 1][22]
Some farming ant species gather and store the aphid eggs in their nests over the winter. In the spring, the ants carry the newly hatched aphids back to the plants. Some species of dairying ants (such as the European yellow meadow ant, Lasius flavus)[23] manage large herds of aphids that feed on roots of plants in the ant colony. Queens leaving to start a new colony take an aphid egg to found a new herd of underground aphids in the new colony. These farming ants protect the aphids by fighting off aphid predators.[22]
An interesting variation in ant–aphid relationships involves lycaenid butterflies and Myrmica ants. For example, Niphanda fusca butterflies lay eggs on plants where ants tend herds of aphids. The eggs hatch as caterpillars which feed on the aphids. The ants do not defend the aphids from the caterpillars (this is due to the caterpillar producing a pheromone the ants detect making them think the caterpillar is actually one of them), but carry the caterpillars to their nest. In the nest, the ants feed the caterpillars, who in return produce honeydew for the ants. When the caterpillars reach full size, they crawl to the colony entrance and form cocoons. After two weeks, butterflies emerge and take flight. At this point the ants will attack the butterfly but the butterfly has a sticky wool like substance on their wings that disable the ants jaws meaning it can take flight without being ripped apart by the ants.[24]:78–79
Some bees in coniferous forests also collect aphid honeydew to make forest honey.[8]
Bacterial endosymbiosis
Endosymbiosis with micro-organisms is common in insects, with more than 10% of insect species relying upon intracellular bacteria for their development and survival [25] Aphids harbour a vertically transmitted (from parent to its offspring) obligate symbiosis with Buchnera aphidicola (Buchner) (Proteobacteria:Enterobacteriaceae), referred to as the primary symbiont, which is located inside specialised cells, the bacteriocytes.[26] The original contamination occurred in a common ancestor 280 to 160 million years ago and has enabled aphids to exploit a new ecological niche, phloem-sap feeding on vascular plants. B. aphidicola provides its host with essential amino acids, which are present in low concentrations in plant sap. The stable intracellular conditions, as well as the bottleneck effect experienced during the transmission of a few bacteria from the mother to each nymph, increase the probability of transmission of mutations and gene deletions.[27][28] As a result, the size of the B. aphidicola genome is greatly reduced, compared to its putative ancestor.[29] Despite the apparent loss of transcription factors in the reduced genome, gene expression is highly regulated, as shown by the ten-fold variation in expression levels between different genes under normal conditions.[30] Buchnera aphidicola gene transcription, although not well understood, is thought to be regulated by a small number of global transcriptional regulators and/or through nutrient supplies from the aphid host.
Some aphid colonies also harbour other bacterial symbionts, referred to as secondary symbionts due to their facultative status. They are vertically transmitted, although some studies demonstrated the possibility of horizontal transmission (from one lineage to another and possibly from one species to another).[31][32] So far, the role of only some of the secondary symbionts has been described; Regiella insecticola plays a role in defining the host-plant range,[33][34] Hamiltonella defensa provides resistance to parasitoids,[35] and Serratia symbiotica prevents the deleterious effects of heat.[36]
Carotenoid synthesis
Some species of aphids have acquired the ability to synthesise red carotenoids by horizontal gene transfer from fungi.[37] This allows otherwise green aphids to be coloured red. Other than the two-spotted spider mites[38] aphids are the only known member of the animal kingdom with the ability to synthesise carotenoids.
Carotenoids may absorb solar energy and convert it to ATP, the first example of photoheterotrophy in animals. The carotene pigments in aphids form a layer close to the surface of the cuticle, where it is ideally placed to absorb sunlight. The excited carotenoids seem to reduce NAD to NADH which can then be oxidized in the mitochondria for energy. It is unclear why aphids should find it necessary to develop this source of energy when their diet provides them with an excess of sugars.[39]
Reproduction


Some aphid species have unusual and complex reproductive adaptations, while others have fairly simple reproduction. Adaptations include having both sexual and asexual reproduction, creation of eggs or live nymphs, and switches between woody and herbaceous types of host plants at different times of the year.[Note 2]
When a sophisticated reproductive strategy is used, only females are present in the population at the beginning of the seasonal cycle (although a few species of aphids have been found to have both male and female sexes). The overwintering eggs that hatch in the spring result in females, called fundatrices. Reproduction is typically parthenogenetic and viviparous. Eggs are parthenogenetically produced without meiosis[40][41] and the offspring are clonal to their mother. The embryos develop within the mothers' ovarioles, which then give live birth to first-instar female nymphs (viviparous). The offspring resemble their parents in every way except size, and are called virginoparae.
This process iterates throughout the summer, producing multiple generations that typically live 20 to 40 days. Thus, one female hatched in spring may produce thousands of descendants. For example, some species of cabbage aphids (like Brevicoryne brassicae) can produce up to 41 generations of females.
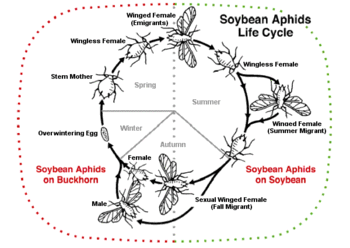
In autumn, aphids undergo sexual, oviparous reproduction. A change in photoperiod and temperature, or perhaps a lower food quantity or quality, causes females to parthenogenetically produce sexual females and males. The males are genetically identical to their mothers except they have one fewer sex chromosome.[40] These sexual aphids may lack wings or even mouthparts.[1] Sexual females and males mate, and females lay eggs that develop outside the mother. The eggs endure the winter and emerge as winged or wingless females the following spring. This is, for example, the lifecycle of the rose aphid (Macrosiphum rosae, or less commonly Aphis rosae), which may be considered typical of the family. However, in warm environments, such as in the tropics or in a greenhouse, aphids may go on reproducing asexually for many years.[8]
Some species produce winged females in the summer, sometimes in response to low food quality or quantity. The winged females migrate to start new colonies on a new plant, often of quite a different kind. For example, the apple aphid (Aphis pomi), after producing many generations of wingless females on its typical food plant, gives rise to winged forms which fly away and settle on grass or corn stalks.
Some aphids have telescoping generations, that is, the parthenogenetic, viviparous female has a daughter within her, who is already parthenogenetically producing her own daughter. Thus, a female's diet can affect the body size and birth rate of more than two generations (daughters and granddaughters).[42][43]
Aphid reproduction terminology:
Heteroecious – host alternating
- Egg
- Fundatrix (foundress from the first egg)
- Fundatrigeniae (daughter clones)
- Emigrant (winged female; in spring, winged aphids migrating from primary hosts infest Poaceae)
- Apterous exule (wingless female)
- Alate exule (winged female)
- Gynoparae (produce sexual females)
- Male
- Oviparae (sexual females that mate with the males)
Autoecious – single host
- Egg
- Fundatrix
- Apterous exule
- Alate exule
- Sexuparae (Parthenogenetic viviparous females of aphids giving rise to the sexual generation and usually developing on the secondary host, the alate forms migrating to the primary host at the end of the summer (holocyclic and heteroecious aphids).)
- Oviparae
- Males

Within these two host lifecycles are other forms: holocyclic (sex involved, will lead to egg production which facilitates overwintering), anholocyclic' (no sex or egg involved, reproduce parthenogenetically), and androcyclic (reproduction at end of season by parthenogenesis to produce males to contribute to holocyclic phase).
The bird cherry-oat aphid is an example of a host-alternating species (as implied by the double name), that starts its lifecycle with a large, highly fecund fundatrix. Her offspring then proceed to grow and produce emigrants which develop on the bird cherry before flying to the oat species where they continue feeding. The subsequent apterous exules feed solely on the oats and eventually lead to growth of gynoparae which will return to the bird cherry, where they will produce males and oviparae, which in turn will reproduce, giving eggs for the next year.
In heteroecious species, the aphids spend winter on tree or bush primary hosts; in summer, they migrate to their secondary host on a herbaceous plant, then the gynoparae return to the tree in autumn. The pea aphid has a primary host of a perennial vetch and secondary of the annual pea. This is likely due to the decline of food quality in trees during the summer, as well as overcrowding amongst aphids which they sense when they bump into each other too often. The heteroecious lifecycle (which is mainly linked to consumption of angiosperms and represents 10% of all aphids) is believed to have evolved from the ancestral autoecious form (on conifers); this is believed to have reverted to the ancestral form in some species that were once heteroecious.
Four types of alate (winged) aphid morphs exist, known as polymorphisms:
- Emigrants (heteroecious only) are produced on primary hosts and migrate to secondary hosts; this is once again due to quality of food decreasing and to a lesser extent overcrowding. These aphids are capable of eating both hosts.
- Alate exules are produced on secondary host if heteroecious, if autoecious will be produced on host anyway. For the alate exules the same factors apply as for emigrants EXCEPT that crowding is more important.
- Gynoparae (heteroecious only and produced on secondary host in response to longer nights and falling temperature). Nymphs can only feed on secondary hosts, and are unable to consume the primary host.
- Males are produced on secondary hosts in heteroecious and in autoecious, normal hosts. These, too, are produced in response to longer nights and decreased temperature. Of these, only 0.6% of autumn alate migrants find host plants, i.e. gynoparae.
Reasons aphids alternate hosts:
- Nutritional optimization (right)
- Temperature tolerance – morphs adapted to part temperature
- Oviposition and rendezvous sites
- Induced host-plant defenses - plants abscise galled tissue; evidence shows that some plants selectively drop galled leaves earlier than ungalled ones.
- Increasing chance of new clones produced
- Autoecious (increase likelihood to meet same individual)
- Heteroecious (decreases chance of meeting self therefore mating with different clone) - better oviposition sites on trees than herbaceous plants as herbaceous plants are annual and die in winter. Problem: survival rate of autoecious vs heteroecious is similar
- Enemy escape – using the same host plants all year round increases the risk of predators discovering the aphids. However, if alates move to other hosts, they and their offspring can circumvent them for a time. One of the problems with this is the individual plants hosting the wingless aphids that have been "left behind" will have large numbers of predators which have discovered them and feed on them.
- Fundatrix specialisation – host alternation is a constraint imposed by specialized feeding requirements of the fundatrix morph as the heteroecious lifecycle is not the optimal one.
- Many host-alternating species are the biggest aphid pests:
- Aphis fabae
- Metopolophium dirhodum
- Myzus persicae
- Rhopalosiphum padi
.JPG)
Evolution
Aphids probably appeared around 280 million years ago, in the early Permian period. They probably fed on plants like Cordaitales or Cycadophyta. The oldest known aphid fossil is of the species Triassoaphis cubitus from the Triassic.[44] The number of species was small, but increased considerably with the appearance of angiosperms 160 million years ago. Angiosperms allowed aphids to specialise. Organs like the cornicles did not appear until the Cretaceous period.
Threats
Aphids are soft-bodied, and have a wide variety of insect predators. Aphids also are often infected by bacteria, viruses, and fungi. They are affected by the weather, such as precipitation,[45] temperature[46] and wind.[47]
Insects that attack aphids include predatory Coccinellidae (lady bugs or ladybirds), hoverfly larvae (Diptera: Syrphidae), parasitic wasps, aphid midge larvae, "aphid lions" (the larvae of green lacewings), and lacewings (Neuroptera: Chrysopidae), and arachnids such as crab spiders.



Fungi that attack aphids include Neozygites fresenii, Entomophthora, Beauveria bassiana, Metarhizium anisopliae, and entomopathogenic fungi such as Lecanicillium lecanii. Aphids brush against the microscopic spores. These spores stick to the aphid, germinate, and penetrate the aphid's skin. The fungus grows in the aphid hemolymph (i.e., the counterpart of blood for aphids). After about 3 days, the aphid dies and the fungus releases more spores into the air. Infected aphids are covered with a woolly mass that progressively grows thicker until the aphid is obscured. Often, the visible fungus is not the type of fungus that killed the aphid, but a secondary fungus.[45]
Aphids can be easily killed by unfavourable weather, such as late spring freezes.[48] Excessive heat kills the symbiotic bacteria that some aphids depend on, which makes the aphids infertile.[49] Rain prevents winged aphids from dispersing, and knocks aphids off plants and thus kills them from the impact or by starvation.[45][50][51] However, rain cannot be relied on for aphid control.[52]
Defenses

Aphids have little protection from predators and diseases. Some species interact with plant tissues forming a gall, an abnormal swelling of plant tissue. Aphids can live inside the gall, which provides protection from predators and the elements. A number of galling aphid species are known to produce specialised "soldier" forms, sterile nymphs with defensive features which defend the gall from invasion.[8][53][54] For example, Alexander's horned aphids are a type of soldier aphid that has a hard exoskeleton and pincer-like mouthparts.[24]:144 The soldiers of gall forming aphids also carry out the job of cleaning the gall. The honeydew secreted by the aphids is coated in a powdery wax to form "liquid marbles"[55] that the soldiers roll out of the gall through small orifices.[54] Aphids that form closed galls use the plant's vascular system for their plumbing: the inner surfaces of the galls are highly absorbent and wastes are absorbed and carried away by the plant.[54]
Infestation of a variety of Chinese trees by Chinese sumac aphids (Melaphis chinensis) can create a "Chinese gall" which is valued as a commercial product. As "Galla Chinensis", Chinese galls are used in Chinese medicine to treat coughs, diarrhoea, night sweats, dysentery and to stop intestinal and uterine bleeding. Chinese galls are also an important source of tannins.[8]
Though aphids cannot fly for most of their life cycle, they can escape predators and accidental ingestion by herbivores by dropping off the plant they are on.[56]
Some species of aphid, known as "woolly aphids" (Eriosomatinae), excrete a "fluffy wax coating" for protection.[8]
The cabbage aphid, Brevicoryne brassicae, stores and releases chemicals that produce a violent chemical reaction and strong mustard oil smell to repel predators.
It was common at one time to suggest that the cornicles were the source of the honeydew, and this was even included in the Shorter Oxford English Dictionary[57] and the 2008 edition of the World Book Encyclopedia.[58] In fact, honeydew secretions are produced from the anus of the aphid,[59] while cornicles mostly produce defensive chemicals such as waxes. There also is evidence of cornicle wax attracting aphid predators in some cases.[60] Aphids are also known to defend themselves from attack by parasitoid wasps by kicking.
Effects on plants

Plants exhibiting aphid damage can have a variety of symptoms, such as decreased growth rates, mottled leaves, yellowing, stunted growth, curled leaves, browning, wilting, low yields and death. The removal of sap creates a lack of vigour in the plant, and aphid saliva is toxic to plants. Aphids frequently transmit disease-causing organisms like plant viruses to their hosts. The green peach aphid, Myzus persicae, is a vector for more than 110 plant viruses. Cotton aphids (Aphis gossypii) often infect sugarcane, papaya and peanuts with viruses.[1] Aphids contributed to the spread of late blight (Phytophthora infestans) among potatoes in the Irish potato famine of the 1840s.[61]
The cherry aphid or black cherry aphid, Myzus cerasi, is responsible for some leaf curl of cherry trees. This can easily be distinguished from 'leaf curl' caused by Taphrina fungus species due to the presence of aphids beneath the leaves.
In plants which produce the phytoestrogen coumestrol, such as alfalfa, damage by aphids is linked with higher concentrations of coumestrol.[62]

The coating of plants with honeydew can contribute to the spread of fungi which can damage plants.[63][64] Honeydew produced by aphids has been observed to reduce the effectiveness of fungicides as well.[65]
A hypothesis that insect feeding may improve plant fitness was floated in the mid-1970s by Owen and Wiegert. It was felt that the excess honeydew would nourish soil micro-organisms, including nitrogen fixers. In a nitrogen poor environment, this could provide an advantage to an infested plant over a noninfested plant. However, this does not appear to be supported by the observational evidence.[66]
The damage of plants, and in particular commercial crops, has resulted in large amounts of resources and efforts being spent attempting to control the activities of aphids.[1]
Some species of aphids of the genus Cinara feed on spruce and fir in North America, but do not cause noticeable injury (Rose and Lindquist 1985).[67] Their long feeding tubes pierce the bark to take up sap from shoots, twigs, branches, stems, and roots. Aphids of most species feed in groups and are usually attended by ants, which feed on the droplets of excreted liquid. The aphids range in colour from grey to brown or black and are less than 5 mm long. All aphids overwinter in the egg stage. Eggs are blackish and are laid singly or in rows on the needles. Six generations in 1 year are not unusual in Canada, with succeeding generations often moving to new sites on the tree, including the roots, as the season progresses. The life cycle is complex. For example, adults of the intermediate summer generations consist of females only, some winged and others wingless, which produce tiny nymphs rather than eggs. Males occur only in the late fall generation, which produces the overwintering eggs.
Control
There are various insecticides that can be used to control aphids, including synthetic insecticides and plant extracts/products that are thought to be more eco-friendly. For example, Shreth et al. suggested use of neem and lantana products to protect plants against aphids.[68]
For small backyard infestations, simply spraying the plants thoroughly with a strong water jet every few days may be sufficient protection for roses and other plants. An insecticidal soap solution can be an effective household remedy to control aphids and other soft-bodied arthropods.[69][70][71] It will only kill aphids on contact and has no residual action against aphids that arrive after application. Soap spray may damage plants, especially at higher concentrations or at temperatures above 32 °C (90 °F). Some plant species are known to be sensitive to soap sprays.
Integrated pest management of various species of aphids can be achieved using biological insecticides based on fungi such as Lecanicillium lecanii, Beauveria bassiana or Paecilomyces fumosoroseus.
Aphids may also be controlled by the release of natural enemies, in particular lady beetles and parasitic wasps. However, since adult lady beetles tend to fly away within 48 hours after release, without laying eggs, it requires repeated application of large numbers of lady beetles to be effective. For example, one large, heavily infested rose bush may take two applications of 1500 beetles each.[69][72] In reality the only cost effective situation in which mass release of natural enemies makes sense is in closed or semi closed environments such as glasshouses or polytunnels.
See also
Notes
References
- 1 2 3 4 5 6 7 8 9 10 George C. McGavin (1993). Bugs of the World. Infobase Publishing. ISBN 0-8160-2737-4.
- ↑ Ross Piper (2007). Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals. Greenwood Press. pp. 6–9. ISBN 978-0-313-33922-6.
- 1 2 http://aphid.speciesfile.org/Common/basic/Taxa.aspx?TaxonNameID=1159459
- ↑ Pip Courtney (October 30, 2005). "Scientist battles lettuce aphid". Landline. Australian Broadcasting Corporation. Retrieved January 1, 2007.
- ↑ R. L. Blackman; V. F. Eastrop (1994). Aphids on the World's Trees. An Identification and Information Guide. Wallingford: CAB International. ISBN 0-85198-877-6.
- 1 2 A. F. G. Dixon (1998). Aphid Ecology (2nd ed.). Chapman and Hall. ISBN 0-412-74180-6.
- ↑ Jeffrey Granett; M. Andrew Walker; Laszlo Kocsis; Amir D. Omer (2001). "Biology and management of grape phylloxera". Annual Review of Entomology. 46: 387–412. doi:10.1146/annurev.ento.46.1.387.
- 1 2 3 4 5 6 7 8 Henry G. Stroyan (1997). "Aphid". McGraw-Hill Encyclopedia of Science and Technology (8th ed.). ISBN 0-07-911504-7.
- ↑ Navdeep S. Mutti (2006). Molecular Studies of the Salivary Glands of the Pea Aphid, Acyrthosiphon pisum (Harris) (PDF) (Ph.D. thesis). Kansas State University.
- ↑ N. J. Spiller; L. Koenders; W. F. Tjallingii (1990). "Xylem ingestion by aphids – a strategy for maintaining water balance". Entomologia Experimentalis et Applicata. 55 (2): 101–104. doi:10.1007/BF00352570.
- 1 2 D. B. Fisher (2000). "Long distance transport". In Bob B. Buchanan; Wilhelm Gruissem; Russell L. Jones. Biochemistry and Molecular Biology of Plants (4th ed.). Rockville, Maryland: American Society of Plant Physiologists. pp. 730–784. ISBN 978-0-943088-39-6.
- ↑ M. Malone; R. Watson; J. Pritchard (1999). "The spittlebug Philaenus spumarius feeds from mature xylem at the full hydraulic tension of the transpiration stream". New Phytologist. 143 (2): 261–271. doi:10.1046/j.1469-8137.1999.00448.x. JSTOR 2588576.
- ↑ Glen Powell; Jim Hardie (2002). "Xylem ingestion by winged aphids". Entomologia Experimentalis et Applicata. 104 (1): 103–108. doi:10.1023/A:1021234412475.
- 1 2 3 Julien Pompon; Dan Quiring; Philippe Giordanengo; Yvan Pelletier (2010). "Role of xylem consumption on osmoregulation in Macrosiphum euphorbiae (Thomas)" (PDF). Journal of Insect Physiology. 56 (6): 610–615. doi:10.1016/j.jinsphys.2009.12.009. PMID 20036244.
- ↑ D. A. Ashford; W. A. Smith; A. E. Douglas (2000). "Living on a high sugar diet: the fate of sucrose ingested by a phloem-feeding insect, the pea aphid Acyrthosiphon pisum". Journal of Insect Physiology. 46 (3): 335–341. doi:10.1016/S0022-1910(99)00186-9. PMID 12770238.
- ↑ T. L. Wilkinson; D. A. Ashfors; J. Pritchard; A. E. Douglas (1997). "Honeydew sugars and osmoregulation in the pea aphid Acyrthosiphon pisum". Journal of Experimental Biology. 200 (11): 2137–2143. PMID 9320049.
- ↑ A. J. Shakesby; I. S. Wallace; H. V. Isaacs; J. Pritchard; D. M. Roberts; A. E. Douglas (2009). "A water-specific aquaporin involved in aphid osmoregulation". Insect Biochemistry and Molecular Biology. 39 (1): 1–10. doi:10.1016/j.ibmb.2008.08.008. PMID 18983920.
- ↑ R. H. Dadd; T. E. Mittler (1965). "Studies on the artificial feeding of the aphid Myzus persicae (Sulzer) – III. Some major nutritional requirements". Journal of Insect Physiology. 11 (6): 717–743. doi:10.1016/0022-1910(65)90154-X. PMID 5827534.
- ↑ Paul Buchner (1965). Endosymbiosis of animals with plant microorganisms. Interscience. ISBN 0-470-11517-3.
- ↑ L. F. Whitehead; A. E. Douglas (1993). "A metabolic study of Buchnera, the intracellular bacterial symbionts of the pea aphid Acyrthosiphon pisum" (PDF). Journal of General Microbiology. 139 (4): 821–826. doi:10.1099/00221287-139-4-821.
- ↑ Gérard Febvay; Isabelle Liadouze; Josette Guillaud; Guy Bonnot (1995). "Analysis of energetic amino acid metabolism in Acyrthosiphon pisum: a multidimensional approach to amino acid metabolism in aphids". Archives of Insect Biochemistry and Physiology. 29 (1): 45–69. doi:10.1002/arch.940290106.
- 1 2 Linda M. Hooper-Bui (2008). "Ant". World Book Encyclopedia. ISBN 978-0-7166-0108-1.
- ↑ Anthony Wootton (1998). Insects of the World. Blandford. ISBN 0-7137-2366-1.
- 1 2 Insects and Spiders, Time-Life Books, ISBN 0-8094-9687-9
- ↑ Paul Baumann; Nancy Moran; Linda Baumann (2006). "Bacteriocyte-associated endosymbionts of insects". In Martin Dworkin; Stanley Falkow; Eugene Rosenberg; Karl-Heinz Schleifer; Erko Stackebrandt. The Prokaryotes. Volume 1. Springer. pp. 403–438. doi:10.1007/0-387-30741-9_16. ISBN 978-0-387-25476-0.
- ↑ A. E. Douglas (1998). "Nutritional interactions in insect-microbial symbioses: aphids and their symbiotic bacteria Buchnera". Annual Review of Entomology. 43: 17–37. doi:10.1146/annurev.ento.43.1.17. PMID 15012383.
- ↑ Vicente Pérez-Broca; Rosario Gil; Silvia Ramos; Araceli Lamelas; Marina Postigo; José Manuel Michelena; Francisco J. Silva; Andrés Moya; Amparo Latorre (2006). "A small microbial genome: The end of a long symbiotic relationship?". Science. 314 (5797): 312–313. doi:10.1126/science.1130441. PMID 17038625.
- ↑ A. Mira; N. A. Moran (2002). "Estimating population size and transmission bottlenecks in maternally transmitted endosymbiotic bacteria". Microbial Ecology. 44 (2): 137–143. doi:10.1007/s00248-002-0012-9. JSTOR 4287640. PMID 12087426.
- ↑ Shuji Shigenobu; Hidemi Watanabe; Masahira Hattori; Yoshiyuki Sakaki; Hajime Ishikawa (2000). "Genome sequence of the endocellular bacterial symbiont of aphids Buchnera sp. APS". Nature. 407 (6800): 81–86. doi:10.1038/35024074. PMID 10993077.
- ↑ José Viñuelas; Federica Calevro; Didier Remond; Jacques Bernillon; Yvan Rahbé; Gérard Febvay; Jean-Michel Fayard; Hubert Charles (2007). "Conservation of the links between gene transcription and chromosomal organisation in the highly reduced genome of Buchnera aphidicola". BMC Genomics. 8: 143. doi:10.1186/1471-2164-8-143. PMC 1899503
 . PMID 17547756.
. PMID 17547756. - ↑ T. Tsuchida; R. Koga; X. Y. Meng; T. Matsumoto; T. Fukatsu (2005). "Characterization of a facultative endosymbiotic bacterium of the pea aphid Acyrthosiphon pisum". Microbial Ecology. 49 (1): 126–133. doi:10.1007/s00248-004-0216-2. PMID 15690225.
- ↑ Makiko Sakurai; Ryuichi Koga; Tsutomu Tsuchida; Xian-Ying Meng; Takema Fukatsu (2005). "Rickettsia symbiont in the pea aphid Acyrthosiphon pisum: novel cellular tropism, effect on host fitness, and interaction with the essential symbiont Buchnera". Applied and Environmental Microbiology. 71 (7): 4069–4075. doi:10.1128/AEM.71.7.4069-4075.2005. PMC 1168972. PMID 16000822.
- ↑ Julia Ferrari; Claire L. Scarborough; H. Charles J. Godfray (2007). "Genetic variation in the effect of a facultative symbiont on host-plant use by pea aphids". Oecologia. 153 (2): 323–329. doi:10.1007/s00442-007-0730-2. PMID 17415589.
- ↑ J.-C. Simon; S. Carré; M. Boutin; N. Prunier–Leterme; B. Sabater–Muñoz; A. Latorre; R. Bournoville (2003). "Host-based divergence in populations of the pea aphid: insights from nuclear markers and the prevalence of facultative symbionts". Proceedings of the Royal Society B: Biological Sciences. 270 (1525): 1703–1712. doi:10.1098/rspb.2003.2430. PMC 1691435. PMID 12964998.
- ↑ K. M. Oliver; N. A. Moran; M. S. Hunter (2006). "Costs and benefits of a superinfection of facultative symbionts in aphids" (PDF). Proceedings of the Royal Society B: Biological Sciences. 273 (1591): 1273–1280. doi:10.1098/rspb.2005.3436. PMC 1560284. PMID 16720402.
- ↑ Gaelen Burke; Oliver Fiehn; Nancy Moran (2009). "Effects of facultative symbionts and heat stress on the metabolome of pea aphids" (PDF). The ISME Journal. 4 (2): 242–252. doi:10.1038/ismej.2009.114. PMID 19907504.
- ↑ Nancy A. Moran; Tyler Jarvik (2010). "Lateral transfer of genes from fungi underlies carotenoid production in aphids". Science. 328 (5978): 624–627. doi:10.1126/science.1187113. PMID 20431015.
- ↑ Altincicek, B; Kovacs, JL; Gerardo, NM (2012). "Horizontally transferred fungal carotenoid genes in the two-spotted spider mite Tetranychus urticae". Biology Letters. 8 (2): 253–257. doi:10.1098/rsbl.2011.0704. PMC 3297373. PMID 21920958.
- ↑ Valmalette, Jean Christophe; Dombrovsky, Aviv; Brat, Pierre; Mertz, Christian; Capovilla, Maria; Robichon, Alain (2012). "Light- induced electron transfer and ATP synthesis in a carotene synthesizing insect". Scientific Reports. 2. doi:10.1038/srep00579.
- 1 2 Hales, Dinah F.; Wilson, Alex C. C.; Sloane, Mathew A.; Simon, Jean-Christophe; Legallic, Jean-François; Sunnucks, Paul (2002). "Lack of Detectable Genetic Recombination on the X Chromosome During the Parthenogenetic Production of Female and Male Aphids". Genetics Research. 79 (3): 203–209. doi:10.1017/S0016672302005657.
- ↑ Blackman, Roger L (1979-05-01). "Stability and variation in aphid clonal lineages". Biological Journal of the Linnean Society. 11 (3): 259–277. doi:10.1111/j.1095-8312.1979.tb00038.x. ISSN 1095-8312. Retrieved 2011-12-21.
- ↑ Ettay Nevo; Moshe Coll (2001). "Effect of nitrogen fertilization on Aphis gossypii (Homoptera: Aphididae): variation in size, color, and reproduction". Journal of Economic Entomology. 94 (1): 27–32. doi:10.1603/0022-0493-94.1.27. PMID 11233124.
- ↑ Gary C. Jahn; Liberty P. Almazan; Jocelyn B. Pacia (2005). "Effect of nitrogen fertilizer on the intrinsic rate of increase of the rusty plum aphid, Hysteroneura setariae (Thomas) (Homoptera: Aphididae) on rice (Oryza sativa L.)" (PDF). Environmental Entomology. 34 (4): 938–943. doi:10.1603/0046-225X-34.4.938.
- ↑ Christine Johnson; Donat Agosti; Jocques H. Delabie; Klaus Dumpert; D. J. Williams; Michael von Tschirnhaus; Ulrich Macshwitz (2001). "Acropyga and Azteca ants (Hymenoptera: Formicidae) with scale insects (Sternorrhyncha: Coccoidea): 20 million years of intimate symbiosis" (PDF). American Museum Novitates. 3335: 1–18. doi:10.1206/0003-0082(2001)335<0001:AAAAHF>2.0.CO;2. ISSN 0003-0082.
- 1 2 3 Gerald E. Brust (June 22, 2006). "Early season aphid and thrips populations". University of Maryland, College Park. Retrieved October 18, 2010.
- ↑ K. P. Lamb (1961). "Some effects of fluctuating temperatures on metabolism, development, and rate of population growth in the cabbage aphid, Brevicoryne brassicae". Ecology. 42 (4): 740–745. doi:10.2307/1933502. JSTOR 1933502.
- ↑ Margaret G. Jones (1979). "Abundance of aphids on cereals from before 1973 to 1977". Journal of Applied Ecology. 16 (1): 1–22. doi:10.2307/2402724. JSTOR 2402724.
- ↑ Christian Krupke; John Obermeyer; Robert O'Neil (May 11, 2007). "Soybean aphid, a new beginning for 2007". Pest and Crop. Purdue University. 7.
- ↑ "Why some aphids can't stand the heat". Science Daily. April 23, 2007.
- ↑ R. D. Hughes (1963). "Population dynamics of the cabbage aphid, Brevicoryne brassicae (L.)". Journal of Animal Ecology. 32 (3): 393–424. doi:10.2307/2600. JSTOR 2600.
- ↑ S. Suwanbutr (1996). "Stable age distributions of lucerne aphid populations in SE-Tasmania" (PDF). Thammasat International Journal of Science and Technology. 1 (5): 38–43.
- ↑ Ken Ostlie (August 3, 2006). "Spider Mites, Aphids and Rain Complicating Spray Decisions in Soybean" (PDF). Minnesota Crop eNews. University of Minnesota.
- ↑ S. Aoki (1977). "Colophina clematis (Homoptera, Pemphigidae), an aphid species with soldiers" (PDF). Japanese Journal of Entomology. 45 (2): 276–282.
- 1 2 3 Kutsukake, M.; Meng, X. Y.; Katayama, N.; Nikoh, N.; Shibao, H.; Fukatsu, T. (2012). "An insect-induced novel plant phenotype for sustaining social life in a closed system". Nature Communications. 3: 1187–. doi:10.1038/ncomms2187. PMC 3514493. PMID 23149732.
- ↑ Pike, N.; Richard, D.; Foster, W.; Mahadevan, L. (2002). "How aphids lose their marbles". Proceedings of the Royal Society B: Biological Sciences. 269 (1497): 1211. doi:10.1098/rspb.2002.1999.
- ↑ Gish, M.; Dafni, A.; Inbar, M. (2012). Heil, Martin, ed. "Young Aphids Avoid Erroneous Dropping when Evading Mammalian Herbivores by Combining Input from Two Sensory Modalities". PLoS ONE. 7 (4): e32706. doi:10.1371/journal.pone.0032706. PMC 3322135. PMID 22496734.
- ↑ John S. Edwards (1966). "Defence by smear: supercooling in the cornicle wax of aphids". Nature. 211 (5044): 73–74. doi:10.1038/211073a0.
- ↑ Candace Martinson (2008). "Aphid". World Book Encyclopedia. ISBN 978-0-7166-0108-1.
- ↑ M. J. Way (1963). "Mutualism between ants and honeydew-producing Homoptera". Annual Review of Entomology. 8: 307–344. doi:10.1146/annurev.en.08.010163.001515.
- ↑ Tessa R. Grasswitz; Timothy D. Paine (1992). "Kairomonal effect of an aphid cornicle secretion on Lysiphlebus testaceipes (Cresson) (Hymenoptera: Aphidiidae)". Journal of Insect Behavior. 5 (4): 447–457. doi:10.1007/BF01058190.
- ↑ Catherine Nichols (2007). The Most Extreme Bugs. John Wiley and Sons. p. 61. ISBN 978-0-7879-8663-6.
- ↑ United States Department of Agriculture (2003). Studies on the Chemical and Biological Properties of Coumestrol and Related Compounds. US Government Printing Office. pp. 47–67.
- ↑ Daniel H. Gillman (2005). "Sooty mold" (PDF). University of Massachusetts Amherst. Retrieved October 18, 2010.
- ↑ Hannah T. Reynolds; Tom Volk (September 2007). "Scorias spongiosa, the beech aphid poop-eater". Tom Volk's Fungus of the Month. University of Wisconsin–La Crosse. Retrieved October 18, 2010.
- ↑ A. J. Dik; J. A. van Pelt (1992). "Interaction between phyllosphere yeasts, aphid honeydew and fungicide effectiveness in wheat under field conditions". Plant Pathology. 41 (6): 661–675. doi:10.1111/j.1365-3059.1992.tb02550.x.
- ↑ Dhrupad Choudhury (1985). "Aphid honeydew: a re-appraisal of the hypothesis of Owen and Wiegert". Oikos. 45 (2): 287–290. doi:10.2307/3565718. JSTOR 3565718.
- ↑ Rose, A.H.; Lindquist, O.H. 1985. Insects of eastern spruces, fir and, hemlock, revised edition. Government of Canada, Canadian Forestry Service, Ottawa, For. Tech. Rep. 23. 159 pp. (cited in Coates et al. 1994, cited orig ed 1977)
- ↑ Chongtham Narajyot Shreth; Kh. Ibohal; S. John William (2009). "Laboratory Evaluation of Certain Cow Urine Extract of Indigenous Plants Against Mustard Aphid, Lipaphis erysimi (Kaltenbach) Infesting Cabbage". Hexapoda. pp. 11–13.
- 1 2 Aphids: Integrated Pest Management for Home Gardeners and Landscape Professionals. M.L. Flint, University of California Davis, July 2013.
- ↑ Insect Control: Soaps and Detergents. W.S. Cranshaw, University of Colorado. March 2008.
- ↑ Insecticidal Soaps for Garden Pest Control. J. D. Ubl, Clemson University, July 2009.
- ↑ Lady Beetle Releases for Aphid Control: How to Help Them Work. J.K. Clark, University of California Davis, June 2011.
External links
| Look up aphid in Wiktionary, the free dictionary. |
| Wikisource has the text of the 1911 Encyclopædia Britannica article Aphides. |
| Wikimedia Commons has media related to Aphidoidea. |
| External identifiers for Aphidoidea | |
|---|---|
| Encyclopedia of Life | 551 |
| ITIS | 109190 |
| NCBI | 33385 |
| Also found in: Wikispecies, ToLWeb | |
- Aphids of southeastern U.S. woody ornamentals
- Acyrthosiphon pisum, MetaPathogen – facts, life cycle, life cycle image
- Sequenced Genome of Pea Aphid, Agricultural Research Service
- Insect Olfaction of Plant Odour: Colorado Potato Beetle and Aphid Studies
- Asian woolly hackberry aphid, Center for Invasive Species Research
On the University of Florida / Institute of Food and Agricultural Sciences Featured Creatures website:
- Aphis gossypii, melon or cotton aphid
- Aphis nerii, oleander aphid
- Hyadaphis coriandri, coriander aphid
- Longistigma caryae, giant bark aphid
- Myzus persicae, green peach aphid
- Sarucallis kahawaluokalani, crapemyrtle aphid
- Shivaphis celti, an Asian woolly hackberry aphid
- Toxoptera citricida, brown citrus aphid