Ferredoxin
Ferredoxins (from Latin ferrum: iron + redox, often abbreviated "fd") are iron-sulfur proteins that mediate electron transfer in a range of metabolic reactions. The term "ferredoxin" was coined by D.C. Wharton of the DuPont Co. and applied to the "iron protein" first purified in 1962 by Mortenson, Valentine, and Carnahan from the anaerobic bacterium Clostridium pasteurianum.[1][2]
Another redox protein, isolated from spinach chloroplasts, was termed "chloroplast ferredoxin".[3] The chloroplast ferredoxin is involved in both cyclic and non-cyclic photophosphorylation reactions of photosynthesis. In non-cyclic photophosphorylation, ferredoxin is the last electron acceptor thus reducing the enzyme NADP+ reductase. It accepts electrons produced from sunlight-excited chlorophyll and transfers them to the enzyme ferredoxin: NADP+ oxidoreductase EC 1.18.1.2.
Ferredoxins are small proteins containing iron and sulfur atoms organized as iron-sulfur clusters. These biological "capacitors" can accept or discharge electrons, with the effect of a change in the oxidation state of the iron atoms between +2 and +3. In this way, ferredoxin acts as an electron transfer agent in biological redox reactions.
Other bioinorganic electron transport systems include rubredoxins, cytochromes, blue copper proteins, and the structurally related Rieske proteins.
Ferredoxins can be classified according to the nature of their iron-sulfur clusters and by sequence similarity.
Fe2S2 ferredoxins
| 2Fe-2S iron-sulfur cluster binding domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
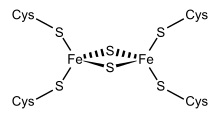 Structural representation of an Fe2S2 ferredoxin. | |||||||||
| Identifiers | |||||||||
| Symbol | Fer2 | ||||||||
| Pfam | PF00111 | ||||||||
| InterPro | IPR001041 | ||||||||
| PROSITE | PDOC00642 | ||||||||
| SCOP | 3fxc | ||||||||
| SUPERFAMILY | 3fxc | ||||||||
| OPM protein | 1kf6 | ||||||||
| |||||||||
Members of the 2Fe-2S ferredoxin family have a general core structure consisting of beta(2)-alpha-beta(2), which includes putidaredoxin, terpredoxin, and adrenodoxin.[4][5][6][7] They are proteins of around one hundred amino acids with four conserved cysteine residues to which the 2Fe-2S cluster is ligated. This conserved region is also found as a domain in various metabolic enzymes and in multidomain proteins, such as aldehyde oxidoreductase (N-terminal), xanthine oxidase (N-terminal), phthalate dioxygenase reductase (C-terminal), succinate dehydrogenase iron-sulphur protein (N-terminal), and methane monooxygenase reductase (N-terminal).
Plant-type ferredoxins
One group of ferredoxins, originally found in chloroplast membranes, has been termed "chloroplast-type" or "plant-type". Its active center is a [Fe2S2] cluster, where the iron atoms are tetrahedrally coordinated both by inorganic sulfur atoms and by sulfurs of four conserved cysteine (Cys) residues.
In chloroplasts, Fe2S2 ferredoxins function as electron carriers in the photosynthetic electron transport chain and as electron donors to various cellular proteins, such as glutamate synthase, nitrite reductase and sulfite reductase. In hydroxylating bacterial dioxygenase systems, they serve as intermediate electron-transfer carriers between reductase flavoproteins and oxygenase.
Thioredoxin-like ferredoxins
The Fe2S2 ferredoxin from Clostridium pasteurianum (Cp2FeFd) has been recognized as distinct protein family on the basis of its amino acid sequence, spectroscopic properties of its iron-sulfur cluster and the unique ligand swapping ability of two cysteine ligands to the [Fe2S2] cluster. Although the physiological role of this ferredoxin remains unclear, a strong and specific interaction of Cp2FeFd with the molybdenum-iron protein of nitrogenase has been revealed. Homologous ferredoxins from Azotobacter vinelandii (Av2FeFdI) and Aquifex aeolicus (AaFd) have been characterized. The crystal structure of AaFd has been solved. AaFd exists as a dimer. The structure of AaFd monomer is different from other Fe2S2 ferredoxins. The fold belongs to the α+β class, with first four β-strands and two α-helices adopting a variant of the thioredoxin fold.
Adrenodoxin-type ferredoxins
| ferredoxin 1 | |
|---|---|
 Crystal structure of human ferredoxin-1 (FDX1).[8] | |
| Identifiers | |
| Symbol | FDX1 |
| Alt. symbols | FDX |
| Entrez | 2230 |
| HUGO | 3638 |
| OMIM | 103260 |
| RefSeq | NM_004109 |
| UniProt | P10109 |
| Other data | |
| Locus | Chr. 11 q22.3 |
Adrenodoxin (adrenal ferredoxin) is expressed in mammals including humans. The human variant of adrenodoxin is referred to as ferredoxin 1. Adrenodoxin, putidaredoxin, and terpredoxin are soluble Fe2S2 proteins that act as single electron carriers. In mitochondrial monooxygenase systems, adrenodoxin transfers an electron from NADPH:adrenodoxin reductase to membrane-bound cytochrome P450. In bacteria, putidaredoxin and terpredoxin serve as electron carriers between corresponding NADH-dependent ferredoxin reductases and soluble P450s. The exact functions of other members of this family are not known, although Escherichia coli Fdx is shown to be involved in biogenesis of Fe-S clusters. Despite low sequence similarity between adrenodoxin-type and plant-type ferredoxins, the two classes have a similar folding topology.
Ferredoxin-1 in humans participates in the synthesis of thyroid hormones. It also transfers electrons from adrenodoxin reductase to the cholesterol side chain cleavage cytochrome P450. FDX-1 has the capability to bind to metals and proteins. It can be found within the cellular mitochondrial matrix.
Fe4S4 and Fe3S4 ferredoxins
The [Fe4S4] ferredoxins may be further subdivided into low-potential (bacterial-type) and high-potential (HiPIP) ferredoxins.
Low- and high-potential ferredoxins are related by the following redox scheme:
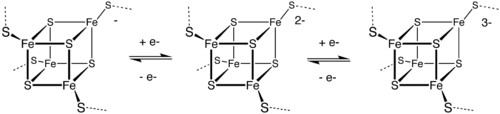
The formal oxidation numbers of the iron ions can be [2Fe3+, 2Fe2+] or [1Fe3+, 3Fe2+] in low-potential ferredoxins. The oxidation numbers of the iron ions in high-potential ferredoxins can be [3Fe3+, 1Fe2+] or [2Fe3+, 2Fe2+].
Bacterial-type ferredoxins
| 3Fe-4S binding domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
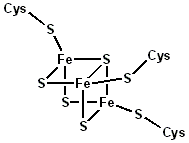 Structural representation of an Fe3S4 ferredoxin. | |||||||||
| Identifiers | |||||||||
| Symbol | Fer4 | ||||||||
| Pfam | PF00037 | ||||||||
| InterPro | IPR001450 | ||||||||
| PROSITE | PDOC00176 | ||||||||
| SCOP | 5fd1 | ||||||||
| SUPERFAMILY | 5fd1 | ||||||||
| OPM protein | 1kqf | ||||||||
| |||||||||
A group of Fe4S4 ferredoxins, originally found in bacteria, has been termed "bacterial-type". Bacterial-type ferredoxins may in turn be subdivided into further groups, based on their sequence properties. Most contain at least one conserved domain, including four cysteine residues that bind to a [Fe4S4] cluster. In Pyrococcus furiosus Fe4S4 ferredoxin, one of the conserved Cys residues is substituted with aspartic acid.
During the evolution of bacterial-type ferredoxins, intrasequence gene duplication, transposition and fusion events occurred, resulting in the appearance of proteins with multiple iron-sulfur centers. In some bacterial ferredoxins, one of the duplicated domains has lost one or more of the four conserved Cys residues. These domains have either lost their iron-sulfur binding property or bind to a [Fe3S4] cluster instead of a [Fe4S4] cluster[9] and dicluster-type.[10]
3-D structures are known for a number of monocluster and dicluster bacterial-type ferredoxins. The fold belongs to the α+β class, with 2-7 α-helices and four β-strands forming a barrel-like structure, and an extruded loop containing three "proximal" Cys ligands of the iron-sulfur cluster.
High-potential iron-sulfur proteins
High-potential iron-sulfur proteins (HiPIPs) form a unique family of Fe4S4 ferredoxins that function in anaerobic electron transport chains. Some HiPIPs have a redox potential higher than any other known iron-sulfur protein (e.g., HiPIP from Rhodopila globiformis has a redox potential of ca. 450 mV). Several HiPIPs have so far been characterized structurally, their folds belonging to the α+β class. As in other bacterial ferredoxins, the [Fe4S4] unit forms a cubane-type cluster and is ligated to the protein via four Cys residues.
Human proteins from ferredoxin family
References
- ↑ Mortenson LE, Valentine RC, Carnahan JE (June 1962). "An electron transport factor from Clostridium pasteurianum". Biochem. Biophys. Res. Commun. 7: 448–52. doi:10.1016/0006-291X(62)90333-9. PMID 14476372.
- ↑ Valentine RC (December 1964). "Bacterial ferredoxin". Bacteriol Rev. 28: 497–517. PMC 441251
 . PMID 14244728.
. PMID 14244728. - ↑ Tagawa K, Arnon DI (August 1962). "Ferredoxins as electron carriers in photosynthesis and in the biological production and consumption of hydrogen gas". Nature. 195 (4841): 537–43. Bibcode:1962Natur.195..537T. doi:10.1038/195537a0. PMID 14039612.
- ↑ Jouanneau Y, Armengaud J, Sainz G, Sieker LC (2001). "Crystallization and preliminary X-ray diffraction analysis of a [2Fe-2S] ferredoxin (FdVI) from Rhodobacter capsulatus". Acta Crystallogr. D. 57 (Pt 2): 301–303. doi:10.1107/S0907444900017832. PMID 11173487.
- ↑ Sevrioukova IF (2005). "Redox-dependent Structural Reorganization in Putidaredoxin, a Vertebrate-type [2Fe-2S] Ferredoxin from Pseudomonas putida". J. Mol. Biol. 347 (3): 607–621. doi:10.1016/j.jmb.2005.01.047. PMID 15755454.
- ↑ Pochapsky TC, Mo H, Pochapsky SS (1999). "A model for the solution structure of oxidized terpredoxin, a Fe2S2 ferredoxin from Pseudomonas". Biochemistry. 38 (17): 5666–5675. doi:10.1021/bi983063r. PMID 10220356.
- ↑ Ruterjans H, Beilke D, Weiss R, Lohr F, Pristovsek P, Hannemann F, Bernhardt R (2002). "A new electron transport mechanism in mitochondrial steroid hydroxylase systems based on structural changes upon the reduction of adrenodoxin". Biochemistry. 41 (25): 7969–7978. doi:10.1021/bi0160361. PMID 12069587.
- ↑ PDB: 3P1M; Chaikuad A, Johansson, C, Krojer, T, Yue, WW, Phillips, C, Bray, JE, Pike, ACW, Muniz, JRC, Vollmar, M, Weigelt, J, Arrowsmith, CH, Edwards, AM, Bountra, C, Kavanagh, K, Oppermann, U (2010). "Crystal structure of human ferredoxin-1 (FDX1) in complex with iron-sulfur cluster". To be published. doi:10.2210/pdb3p1m/pdb.
- ↑ Fukuyama K, Matsubara H, Katsube Y, Tsukihara T (1989). "Structure of [4Fe-4S] ferredoxin from Bacillus thermoproteolyticus refined at 2.3 A resolution. Structural comparisons of bacterial ferredoxins". J. Mol. Biol. 210 (2): 383–398. doi:10.1016/0022-2836(89)90338-0. PMID 2600971.
- ↑ Sieker LC, Meyer J, Moulis JM, Fanchon E, Duee ED, Vicat J (1994). "Refined crystal structure of the 2[4Fe-4S] ferredoxin from Clostridium acidurici at 1.84 A resolution". J. Mol. Biol. 243 (4): 683–695. doi:10.1016/0022-2836(94)90041-8. PMID 7966291.
Further reading
- Bruschi, M.; Guerlesquin, F. (1988). "Structure, function and evolution of bacterial ferredoxins". FEMS Microbiol. Rev. 4 (2): 155–75. doi:10.1111/j.1574-6968.1988.tb02741.x. PMID 3078742.
- Ciurli, S.; Musiani, F. (2005). "High potential iron-sulfur proteins and their role as soluble electron carriers in bacterial photosynthesis: tale of a discovery". Photosynth. Res. 85 (1): 115–131. doi:10.1007/s11120-004-6556-4. PMID 15977063.
- Fukuyama, K. (2004). "Structure and function of plant-type ferredoxins". Photosynth. Res. 81 (3): 289–301. doi:10.1023/B:PRES.0000036882.19322.0a. PMID 16034533.
- Grinberg, A.V., Hannemann, F., Schiffler, B., Müller, J., Heinemann, U. and Bernhardt, R. (2000). "Adrenodoxin: structure, stability, and electron transfer properties". Proteins. 40 (4): 590–612. doi:10.1002/1097-0134(20000901)40:4<590::AID-PROT50>3.0.CO;2-P. PMID 10899784.
- Holden,H.M., Jacobson, B.L., Hurley, J.K., Tollin, G., Oh, B.H., Skjeldal, L., Chae, Y.K., Cheng, H., Xia, B. and Markley, J.L. (1994). "Structure-function studies of [2Fe-2S] ferredoxins". J. Bioenerg. Biomembr. 26 (1): 67–88. doi:10.1007/BF00763220. PMID 8027024.
- Meyer, J. (2001). "Ferredoxins of the third kind". FEBS Lett. 509 (1): 1–5. doi:10.1016/S0014-5793(01)03049-6. PMID 11734195.
External links
- InterPro: IPR006057 - 2Fe-2S ferredoxin subdomain
- InterPro: IPR001055 - Adrenodoxin
- InterPro: IPR001450 - 4Fe-4S ferredoxin, iron-sulfur binding
- InterPro: IPR000170 - High potential iron-sulfur protein
- PDB: 1F37 - X-ray structure of thioredoxin-like ferredoxin from Aquifex aeolicus (AaFd)