Lipidomics
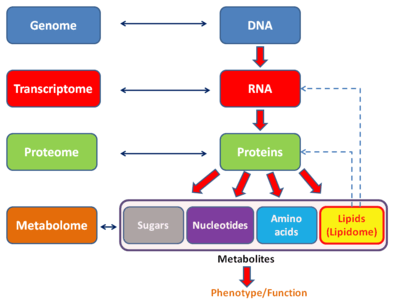
Lipidomics is the large-scale study of pathways and networks of cellular lipids in biological systems[1][2][3] The word "lipidome" is used to describe the complete lipid profile within a cell, tissue, organism, or ecosystem and is a subset of the "metabolome" which also includes the three other major classes of biological molecules: proteins/amino-acids, sugars and nucleic acids. Lipidomics is a relatively recent research field that has been driven by rapid advances in technologies such as mass spectrometry (MS), nuclear magnetic resonance (NMR) spectroscopy, fluorescence spectroscopy, dual polarisation interferometry and computational methods, coupled with the recognition of the role of lipids in many metabolic diseases such as obesity, atherosclerosis, stroke, hypertension and diabetes. This rapidly expanding field[4] complements the huge progress made in genomics and proteomics, all of which constitute the family of systems biology.
Lipidomics research involves the identification and quantification of the thousands of cellular lipid molecular species and their interactions with other lipids, proteins, and other metabolites. Investigators in lipidomics examine the structures, functions, interactions, and dynamics of cellular lipids and the changes that occur during perturbation of the system.
Han and Gross[5] first defined the field of lipidomics through integrating the specific chemical properties inherent in lipid molecular species with a comprehensive mass spectrometric approach. Although lipidomics is under the umbrella of the more general field of "metabolomics", lipidomics is itself a distinct discipline due to the uniqueness and functional specificity of lipids relative to other metabolites.
In lipidomic research, a vast amount of information quantitatively describing the spatial and temporal alterations in the content and composition of different lipid molecular species is accrued after perturbation of a cell through changes in its physiological or pathological state. Information obtained from these studies facilitates mechanistic insights into changes in cellular function. Therefore, lipidomic studies play an essential role in defining the biochemical mechanisms of lipid-related disease processes through identifying alterations in cellular lipid metabolism, trafficking and homeostasis. The growing attention on lipid research is also seen from the initiatives underway of the LIPID Metabolites And Pathways Strategy (LIPID MAPS Consortium).[6] and The European Lipidomics Initiative (ELIfe).[7]

Structural diversity of lipids
Lipids are a diverse and ubiquitous group of compounds which have many key biological functions, such as acting as structural components of cell membranes, serving as energy storage sources and participating in signaling pathways. Lipids may be broadly defined as hydrophobic or amphipathic small molecules that originate entirely or in part from two distinct types of biochemical subunits or "building blocks": ketoacyl and isoprene groups.[8] The huge structural diversity found in lipids arises from the biosynthesis of various combinations of these building blocks. For example, glycerophospholipids are composed of a glycerol backbone linked to one of approximately 10 possible headgroups and also to 2 fatty acyl/alkyl chains, which in turn may have 30 or more different molecular structures. In practice, not all possible permutations are detected experimentally, due to chain preferences depending on the cell type and also to detection limits - nevertheless several hundred distinct glycerophospholipid molecular species have been detected in mammalian cells.
Plant chloroplast thylakoid membranes however, have unique lipid composition as they are deficient in phospholipids. Also, their largest constituent, monogalactosyl diglyceride or MGDG, does not form aqueous bilayers. Nevertheless, dyamic studies reveal a normal lipid bilayer organisation in thylakoid membranes.[9]
Experimental techniques
Lipid extraction
Most methods of lipid extraction and isolation from biological samples exploit the high solubility of hydrocarbon chains in organic solvents. Given the diversity in lipid classes, it is not possible to accommodate all classes with a common extraction method. The traditional Bligh/Dyer procedure [10] uses chloroform/methanol-based protocols that include phase partitioning into the organic layer. These protocols work relatively well for a wide variety of physiologically relevant lipids but they have to be adapted for complex lipid chemistries and low-abundance and labile lipid metabolites.[11][12][13][14][15][16] When organic soil was used, citrate buffer in the extraction mixture gave higher amounts of lipid phosphate than acetate buffer, Tris, H2O or phosphate buffer.[17]
Lipid separation
The simplest method of lipid separation is the use of thin layer chromatography (TLC). Although not as sensitive as other methods of lipid detection, it offers a rapid and comprehensive screening tool prior to more sensitive and sophisticated techniques. Solid-phase extraction (SPE) chromatography is useful for rapid, preparative separation of crude lipid mixtures into different lipid classes. This involves the use of prepacked columns containing silica or other stationary phases to separate glycerophospholipids, fatty acids, cholesteryl esters, glycerolipids, and sterols from crude lipid mixtures.[18] High-performance liquid chromatography (HPLC or LC) is extensively used in lipidomic analysis to separate lipids prior to mass analysis. Separation can be achieved by either normal-phase (NP) HPLC or reverse-phase (RP) HPLC. For example, NP-HPLC effectively separates glycerophospholipids on the basis of headgroup polarity,[19] whereas RP-HPLC effectively separates fatty acids such as eicosanoids on the basis of chain length, degree of unsaturation and substitution.[20] For global, untargeted lipidomic studies it is common to use both RP and NP or Hydrophilic Interaction Liquid Chromatrography (HILC) columns for increased coverage of lipidome. This can be further improved with ultra-performance (UPLC) columns (2.1 mm ID & sub-2 micrometer particle size) or columns based on solid- core technology (2.1 mm ID & 2.6 µm particle size). The UHPLC columns allow high resolution separation of complex lipids with increased peak capacity and sensitivity, whereas solid- core columns achieve same at shorter times on normal HPLC pumps. Chromatographic (HPLC/UHPLC) separation of lipids may either be performed offline or online where the eluate is integrated with the ionization source of a mass spectrometer.
Lipid detection
The progress of modern lipidomics has been greatly accelerated by the development of spectrometric methods in general and soft ionization techniques for mass spectrometry such as electrospray ionization (ESI)[5] and matrix-assisted laser desorption/ionization (MALDI)[21] in particular. "Soft" ionization does not cause extensive fragmentation, so that comprehensive detection of an entire range of lipids within a complex mixture can be correlated to experimental conditions or disease state. In addition to ESI and MALDI, the technique of atmospheric pressure chemical ionization (APCI) has become increasingly popular for the analysis of nonpolar lipids.[22]

ESI MS
ESI-MS was initially developed by Fenn and colleagues for analysis of biomolecules.[23] It depends on the formation of gaseous ions from polar, thermally labile and mostly non-volatile molecules and thus is completely suitable for a variety of lipids. It is a soft-ionization method that rarely disrupts the chemical nature of the analyte prior to mass analysis. Various ESI-MS methods have been developed for analysis of different classes, subclasses, and individual lipid species from biological extracts. Comprehensive reviews of the methods and their application have recently been published.[24] The major advantages of ESI-MS are high accuracy, sensitivity, reproducibility, and the applicability of the technique to complex solutions without prior derivatization. Han and coworkers have developed a method known as"shotgun lipidomics" which involves direct infusion of a crude lipid extract into an ESI source optimized for intrasource separation of lipids based on their intrinsic electrical properties.[25]
MALDI MS
MALDI mass spectrometry is a laser-based soft-ionization method often used for analysis of large proteins, but has been used successfully for lipids. The lipid is mixed with a matrix, such as 2,5-dihydroxybenzoic acid, and applied to a sample holder as a small spot. A laser is fired at the spot, and the matrix absorbs the energy, which is then transferred to the analyte, resulting in ionization of the molecule. MALDI-Time-of-flight (MALDI-TOF) MS has become a very promising approach for lipidomics studies, particularly for the imaging of lipids from tissue slides.[26]
APCI MS
The source for APCI is similar to ESI except that ions are formed by the interaction of the heated analyte solvent with a corona discharge needle set at a high electrical potential. Primary ions are formed immediately surrounding the needle, and these interact with the solvent to form secondary ions that ultimately ionize the sample. APCI is particularly useful for the analysis of nonpolar lipids such as triacylglycerols, sterols, and fatty acid esters.[27]
Imaging techniques
Recent developments in MALDI methods have enabled direct detection of lipids in-situ. Abundant lipid-related ions are produced from the direct analysis of thin tissue slices when sequential spectra are acquired across a tissue surface that has been coated with a MALDI matrix. Collisional activation of the molecular ions can be used to determine the lipid family and often structurally define the molecular species. This technique enables detection of phospholipids, sphingolipids and glycerolipids in tissues such as heart, kidney and brain. Furthermore, distribution of many different lipid molecular species often define anatomical regions within these tissues.[28]
Lipidomic profiling

Lipid profiling is a targeted metabolomics platform that provides a comprehensive analysis of lipid species within a cell or tissue. Profiling based on electrospray ionization tandem mass spectrometry (ESI-MS/MS) is capable of providing quantitative data and is adaptable to high throughput analyses.[30] The powerful approach of transgenics, namely deletion and/or overexpression of a gene product coupled with lipidomics, can give valuable insights into the role of biochemical pathways.[31] Lipid profiling techniques have also been applied to plants[32] and microorganisms such as yeast.[29][33] A combination of quantitative lipidomic data in conjunction with the corresponding transcriptional data (using gene-array methods) and proteomic data (using tandem MS) enables a systems biology approach to a more in-depth understanding of the metabolic or signaling pathways of interest.
Informatics
A major challenge for lipidomics, in particular for MS-based approaches, lies in the computational and bioinformatic demands of handling the large amount of data that arise at various stages along the chain of information acquisition and processing.[34] [35] Chromatographic and MS data collection requires substantial efforts in spectral alignment and statistical evaluation of fluctuations in signal intensities. Such variations have a multitude of origins, including biological variations, sample handling and analytical accuracy. As a consequence several replicates are normally required for reliable determination of lipid levels in complex mixtures. Within the last few years, a number of software packages have been developed by various companies and research groups to analyze data generated by MS profiling of metabolites, including lipids. The data processing for differential profiling usually proceed through several stages, including input file manipulation, spectral filtering, peak detection, chromatographic alignment, normalization, visualization, and data export. An example of metabolic profiling software is the freely-available Java-based Mzmine application.[36] Some software packages such as Markerview[37] include multivariate statistical analysis (for example, principal component analysis) and these will be helpful for the identification of correlations in lipid metabolites that are associated with a physiological phenotype, in particular for the development of lipid-based biomarkers.Another objective of the information technology side of lipidomics involves the construction of metabolic maps from data on lipid structures and lipid-related protein and genes. Some of these lipid pathways[38] are extremely complex, for example the mammalian glycosphingolipid pathway.[39] The establishment of searchable and interactive databases[40][41] of lipids and lipid-related genes/proteins is also an extremely important resource as a reference for the lipidomics community. Integration of these databases with MS and other experimental data, as well as with metabolic networks[42] offers an opportunity to devise therapeutic strategies to prevent or reverse these pathological states involving dysfunction of lipid-related processes.
References
- ↑ Wenk MR (July 2005). "The emerging field of lipidomics". Nat Rev Drug Discov. 4 (7): 594–610. doi:10.1038/nrd1776. PMID 16052242.
- ↑ Watson AD (October 2006). "Thematic review series: systems biology approaches to metabolic and cardiovascular disorders. Lipidomics: a global approach to lipid analysis in biological systems". J. Lipid Res. 47 (10): 2101–11. doi:10.1194/jlr.R600022-JLR200. PMID 16902246.
- ↑ "Lipidomics". The Lipid Chronicles. Retrieved 2012-01-08.
- ↑ Han X (2007). "Neurolipidomics: challenges and developments". Front. Biosci. 12: 2601–15. doi:10.2741/2258. PMC 2141543
 . PMID 17127266.
. PMID 17127266. - 1 2 Han X, Gross RW; Gross (June 2003). "Global analyses of cellular lipidomes directly from crude extracts of biological samples by ESI mass spectrometry: a bridge to lipidomics". J. Lipid Res. 44 (6): 1071–9. doi:10.1194/jlr.R300004-JLR200. PMID 12671038.
- ↑ LIPID MAPS Consortium
- ↑ European Lipidomics Initiative
- ↑ Fahy E, Subramaniam S, Brown HA, et al. (2005). "A comprehensive classification system for lipids". J. Lipid Res. 46 (5): 839–61. doi:10.1194/jlr.E400004-JLR200. PMID 15722563.
- ↑ YashRoy R.C. (1990) Magnetic resonance studies on dynamic organisation of lipids in chloroplast membranes. Journal of Biosciences, vol. 15(4), pp. 281-288.https://www.researchgate.net/publication/225688482_Magnetic_resonance_studies_of_dynamic_organisation_of_lipids_in_chloroplast_membranes?ev=prf_pub
- ↑ Bligh EG, Dyer WJ; Dyer (August 1959). "A rapid method of total lipid extraction and purification". Can J Biochem Physiol. 37 (8): 911–7. doi:10.1139/o59-099. PMID 13671378.
- ↑ Krank J, Murphy RC, Barkley RM, Duchoslav E, McAnoy A; Murphy; Barkley; Duchoslav; McAnoy (2007). "Qualitative analysis and quantitative assessment of changes in neutral glycerol lipid molecular species within cells". Meth. Enzymol. Methods in Enzymology. 432: 1–20. doi:10.1016/S0076-6879(07)32001-6. ISBN 978-0-12-373895-0. PMID 17954211.
- ↑ Ivanova PT, Milne SB, Byrne MO, Xiang Y, Brown HA; Milne; Byrne; Xiang; Brown (2007). "Glycerophospholipid identification and quantitation by electrospray ionization mass spectrometry". Meth. Enzymol. Methods in Enzymology. 432: 21–57. doi:10.1016/S0076-6879(07)32002-8. ISBN 978-0-12-373895-0. PMID 17954212.
- ↑ Deems R, Buczynski MW, Bowers-Gentry R, Harkewicz R, Dennis EA; Buczynski; Bowers-Gentry; Harkewicz; Dennis (2007). "Detection and quantitation of eicosanoids via high performance liquid chromatography-electrospray ionization-mass spectrometry". Meth. Enzymol. Methods in Enzymology. 432: 59–82. doi:10.1016/S0076-6879(07)32003-X. ISBN 978-0-12-373895-0. PMID 17954213.
- ↑ McDonald JG, Thompson BM, McCrum EC, Russell DW; Thompson; McCrum; Russell (2007). "Extraction and analysis of sterols in biological matrices by high performance liquid chromatography electrospray ionization mass spectrometry". Meth. Enzymol. Methods in Enzymology. 432: 145–70. doi:10.1016/S0076-6879(07)32006-5. ISBN 978-0-12-373895-0. PMID 17954216.
- ↑ Garrett TA, Guan Z, Raetz CR; Guan; Raetz (2007). "Analysis of ubiquinones, dolichols, and dolichol diphosphate-oligosaccharides by liquid chromatography-electrospray ionization-mass spectrometry". Meth. Enzymol. Methods in Enzymology. 432: 117–43. doi:10.1016/S0076-6879(07)32005-3. ISBN 978-0-12-373895-0. PMID 17954215.
- ↑ Sullards MC, Allegood JC, Kelly S, Wang E, Haynes CA, Park H, Chen Y, Merrill AH; Allegood; Kelly; Wang; Haynes; Park; Chen; Merrill Jr (2007). "Structure-specific, quantitative methods for analysis of sphingolipids by liquid chromatography-tandem mass spectrometry: "inside-out" sphingolipidomics". Meth. Enzymol. Methods in Enzymology. 432: 83–115. doi:10.1016/S0076-6879(07)32004-1. ISBN 978-0-12-373895-0. PMID 17954214.
- ↑ Å. Frostegård, A. Tunlid & E. Bååth (August 1991). "Microbial biomass measured as total lipid phosphate in soils of different organic content". J. of Microbiological Methods. 14 (3): 151–163. doi:10.1016/0167-7012(91)90018-L.
- ↑ Kaluzny MA, Duncan LA, Merritt MV, Epps DE; Duncan; Merritt; Epps (January 1985). "Rapid separation of lipid classes in high yield and purity using bonded phase columns". J. Lipid Res. 26 (1): 135–40. PMID 3973509.
- ↑ Malavolta M, Bocci F, Boselli E, Frega NG; Bocci; Boselli; Frega (October 2004). "Normal phase liquid chromatography-electrospray ionization tandem mass spectrometry analysis of phospholipid molecular species in blood mononuclear cells: application to cystic fibrosis". J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 810 (2): 173–86. doi:10.1016/j.jchromb.2004.07.001. PMID 15380713.
- ↑ Nakamura T, Bratton DL, Murphy RC; Bratton; Murphy (August 1997). "Analysis of epoxyeicosatrienoic and monohydroxyeicosatetraenoic acids esterified to phospholipids in human red blood cells by electrospray tandem mass spectrometry". J Mass Spectrom. 32 (8): 888–96. doi:10.1002/(SICI)1096-9888(199708)32:8<888::AID-JMS548>3.0.CO;2-W. PMID 9269087.
- ↑ Fuchs B, Schiller J; Schiller (2008). "MALDI-TOF MS analysis of lipids from cells, tissues and body fluids". Subcell. Biochem. Subcellular Biochemistry. 49: 541–65. doi:10.1007/978-1-4020-8831-5_21. ISBN 978-1-4020-8830-8. PMID 18751926.
- ↑ Byrdwell WC (April 2001). "Atmospheric pressure chemical ionization mass spectrometry for analysis of lipids". Lipids. 36 (4): 327–46. doi:10.1007/s11745-001-0725-5. PMID 11383683.
- ↑ Fenn JB, Mann M, Meng CK, Wong SF, Whitehouse CM; Mann; Meng; Wong; Whitehouse (October 1989). "Electrospray ionization for mass spectrometry of large biomolecules". Science. 246 (4926): 64–71. Bibcode:1989Sci...246...64F. doi:10.1126/science.2675315. PMID 2675315.
- ↑ Murphy RC, Fiedler J, Hevko J; Fiedler; Hevko (February 2001). "Analysis of nonvolatile lipids by mass spectrometry". Chem. Rev. 101 (2): 479–526. doi:10.1021/cr9900883. PMID 11712255.
- ↑ Gross RW, Han X; Han (2007). "Lipidomics in diabetes and the metabolic syndrome". Meth. Enzymol. Methods in Enzymology. 433: 73–90. doi:10.1016/S0076-6879(07)33004-8. ISBN 978-0-12-373966-7. PMID 17954229.
- ↑ Schiller J, Suss R, Fuchs B, Muller M, Zschornig O, Arnold K; Suss; Fuchs; Muller; Zschornig; Arnold (2007). "MALDI-TOF MS in lipidomics". Front. Biosci. 12: 2568–79. doi:10.2741/2255. PMID 17127263.
- ↑ Byrdwell WC (2008). "Dual parallel liquid chromatography with dual mass spectrometry (LC2/MS2) for a total lipid analysis". Front. Biosci. 13 (13): 100–20. doi:10.2741/2663. PMID 17981531.
- ↑ Murphy RC, Hankin JA, Barkley RM; Hankin; Barkley (December 2008). "Imaging of lipid species by MALDI mass spectrometry". J. Lipid Res. 50 Suppl (Supplement): S317–22. doi:10.1194/jlr.R800051-JLR200. PMC 2674737. PMID 19050313.
- 1 2 Klose, C; Surma, MA.; Gerl, MJ.; Meyenhofer, F; Shevchenko, A; Simons, K (April 2012). "Flexibility of a Eukaryotic Lipidome – Insights from Yeast Lipidomics". PLoS ONE. 7 (4): e35063. Bibcode:2012PLoSO...7E5063K. doi:10.1371/journal.pone.0035063. PMC 3329542. PMID 22529973.
- ↑ Lipid profiling of a mouse macrophage cell line (LIPID MAPS)
- ↑ Serhan CN, Jain A, Marleau S, Clish C, Kantarci A, Behbehani B, Colgan SP, Stahl GL, Merched A, Petasis NA, Chan L, Van Dyke TE; Jain; Marleau; Clish; Kantarci; Behbehani; Colgan; Stahl; Merched; Petasis; Chan; Van Dyke (December 2003). "Reduced inflammation and tissue damage in transgenic rabbits overexpressing 15-lipoxygenase and endogenous anti-inflammatory lipid mediators". J. Immunol. 171 (12): 6856–65. doi:10.4049/jimmunol.171.12.6856. PMID 14662892.
- ↑ Devaiah SP, Roth MR, Baughman E, Li M, Tamura P, Jeannotte R, Welti R, Wang X; Roth; Baughman; Li; Tamura; Jeannotte; Welti; Wang (September 2006). "Quantitative profiling of polar glycerolipid species from organs of wild-type Arabidopsis and a phospholipase Dalpha1 knockout mutant". Phytochemistry. 67 (17): 1907–24. doi:10.1016/j.phytochem.2006.06.005. PMID 16843506.
- ↑ Ejsing CS, Moehring T, Bahr U, Duchoslav E, Karas M, Simons K, Shevchenko A; Moehring; Bahr; Duchoslav; Karas; Simons; Shevchenko (March 2006). "Collision-induced dissociation pathways of yeast sphingolipids and their molecular profiling in total lipid extracts: a study by quadrupole TOF and linear ion trap-orbitrap mass spectrometry". J Mass Spectrom. 41 (3): 372–89. doi:10.1002/jms.997. PMID 16498600.
- ↑ Subramaniam S, Fahy E, Gupta S, Sud M, Byrnes R.W, Cotter D, Dinasarapu A.R and Maurya M.R (2011). "Bioinformatics and Systems Biology of the Lipidome". Chemical Reviews. 111 (10): 6452–6490. doi:10.1021/cr200295k. PMC 3383319. PMID 21939287.
- ↑ Yetukuri L, Katajamaa M, Medina-Gomez G, Seppänen-Laakso T, Vidal-Puig A, Oresic M; Katajamaa; Medina-Gomez; Seppänen-Laakso; Vidal-Puig; Oresic (2007). "Bioinformatics strategies for lipidomics analysis: characterization of obesity related hepatic steatosis". BMC Syst Biol. 1: 12. doi:10.1186/1752-0509-1-12. PMC 1839890. PMID 17408502.
- ↑ Katajamaa M, Miettinen J, Oresic M; Miettinen; Oresic (March 2006). "MZmine: toolbox for processing and visualization of mass spectrometry based molecular profile data". Bioinformatics. 22 (5): 634–6. doi:10.1093/bioinformatics/btk039. PMID 16403790.
- ↑ Lutz U, Lutz RW, Lutz WK; Lutz; Lutz (July 2006). "Metabolic profiling of glucuronides in human urine by LC-MS/MS and partial least-squares discriminant analysis for classification and prediction of gender". Anal. Chem. 78 (13): 4564–71. doi:10.1021/ac0522299. PMID 16808466.
- ↑ Okuda S, Yamada T, Hamajima M, Itoh M, Katayama T, Bork P, Goto S, Kanehisa M; Yamada; Hamajima; Itoh; Katayama; Bork; Goto; Kanehisa (July 2008). "KEGG Atlas mapping for global analysis of metabolic pathways". Nucleic Acids Res. 36 (Web Server issue): W423–6. doi:10.1093/nar/gkn282. PMC 2447737. PMID 18477636.
- ↑ SphingoMAP
- ↑ Sud M, Fahy E, Cotter D, Brown A, Dennis EA, Glass CK, Merrill AH, Murphy RC, Raetz CR, Russell DW, Subramaniam S; Fahy; Cotter; Brown; Dennis; Glass; Merrill Jr; Murphy; Raetz; Russell; Subramaniam (January 2007). "LMSD: LIPID MAPS structure database". Nucleic Acids Res. 35 (Database issue): D527–32. doi:10.1093/nar/gkl838. PMC 1669719. PMID 17098933.
- ↑ Cotter D, Maer A, Guda C, Saunders B, Subramaniam S; Maer; Guda; Saunders; Subramaniam (January 2006). "LMPD: LIPID MAPS proteome database". Nucleic Acids Res. 34 (Database issue): D507–10. doi:10.1093/nar/gkj122. PMC 1347484. PMID 16381922.
- ↑ Yetukuri L, Ekroos K, Vidal-Puig A, Oresic M; Ekroos; Vidal-Puig; Oresic (February 2008). "Informatics and computational strategies for the study of lipids". Mol Biosyst. 4 (2): 121–7. doi:10.1039/b715468b. PMID 18213405.
External links
| Wikimedia Commons has media related to |