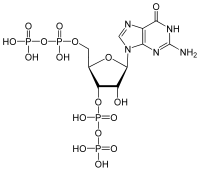
Guanosine pentaphosphate
 | |
| Names | |
|---|---|
| Other names
guanosine pentaphosphate (pppGpp), guanosine tetraphosphate (ppGpp) | |
| Identifiers | |
| 32452-17-8 | |
| DrugBank | DB04022 |
| PubChem | 766 |
| Properties | |
| C10H17N5O17P4 | |
| Molar mass | 603.16 g·mol−1 |
| Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
| Infobox references | |
(p)ppGpp, guanosine pentaphosphate or tetraphosphate is an alarmone which is involved in the stringent response in bacteria, causing the inhibition of RNA synthesis when there is a shortage of amino acids present. This causes translation to decrease and the amino acids present are therefore conserved. Furthermore, ppGpp causes the up-regulation of many other genes involved in stress response such as the genes for amino acid uptake (from surrounding media) and biosynthesis.[1]
Discovery
ppGpp and pppGpp were first identified by Michael Cashel in the 1960s. These nucleotides were found to accumulate rapidly in Escherichia coli cells starved for amino acids, and inhibit synthesis of ribosomal and transfer RNAs.[2] It is now known that (p)ppGpp is also produced in response to other stressors, including carbon and phosphate starvation.
Absence of (p)ppGpp
A complete absence of (p)ppGpp causes multiple amino acid requirements, poor survival of aged cultures, aberrant cell division, morphology, and immotility, as well as being locked in a growth mode during entry into starvation.
Synthesis and degradation of (p)ppGpp
The synthesis and degradation of (p)ppGpp have been most extensively characterized in the model system E. coli. (p)ppGpp is created via pppGpp synthase, also known as RelA, and is converted from pppGpp to ppGpp via pppGpp phosphohydrolase. RelA is associated with about every one in two hundred ribosomes and it becomes activated when an uncharged transfer RNA (tRNA) molecule enters the A site of the ribosome, due to the shortage of amino acid required by the tRNA. If a mutant bacterium is relA− it is said to be relaxed and no regulation of RNA production due to amino acid absence is seen.
E. coli produces a second protein responsible for degradation of (p)ppGpp, termed SpoT. When the amino acid balance in the cell is restored, (p)ppGpp is hydrolysed by SpoT. This protein also has the capacity to synthesize (p)ppGpp, and seems to be the primary synthase under certain conditions of stress. Most other bacteria encode a single protein that is responsible for both synthesis and degradation of (p)ppGpp, generally homologs of SpoT.
Targets of (p)ppGpp
Targets of (p)ppGpp include rRNA operons, of which there are seven in Escherichia coli (a commonly used bacterial model organism), all of which have 2 promoters. When (p)ppGpp associates with the promoter it affects the RNA polymerase enzyme's ability to bind and initiate transcription. It is thought that (p)ppGpp may affect the stability of the open complex formed by RNA polymerase on DNA and therefore affect promoter clearance. Its presence also leads to an increase in pausing during transcription elongation and it competes with nucleoside triphosphate substrates.
There is now a consensus that (p)ppGpp is a determinant of growth rate control rather than nucleoside triphosphate (NTP) substrate concentrations.
Impact of (p)ppGpp on bacterial physiology
Inhibition of growth by inhibition of protein synthesis
ppGpp inhibits IF2-mediated fMet-Phe initiation dipeptide formation, probably by interfering with 30S and 50S subunit interactions. E. coli accumulates more ppGpp than pppGpp during amino acid starvation, and ppGpp has about 8-fold greater efficiency than that of pppGpp. While B. subtilis accumulates more pppGpp than ppGpp.
Inhibition of DNA replication
In E. coli amino acid starvation inhibited DNA replication at the initiation stage at oriC, most probably owing to the lack of the DnaA replication initiation protein. In B. subtilis, the replication arrest due to (p)ppGpp accumulation is caused by the binding of an Rtp protein to specific sites about 100-200kb away from oriC in both directions. DNA primase (DnaG) was directly inhibited by (p)ppGpp. Unlike E. coli, B. subtilis accumulates more pppGpp than ppGpp; the more abundant nucleotide is a more-potent DnaG inhibitor. ppGpp can bind with Obg protein which belongs to the conserved, small GTPase protein family. Obg protein interacts with several regulators (RsbT, RsbW, RsbX) necessary for the stress activation of sigma B.
Impact on phage replication and development
The (p)ppGpp levels of the host seem to act as a sensor for phage lambda development, primarily affecting transcription. Modest ppGpp levels inhibit pR and active pE, pI, and paQ promoters in vivo and have effects in vitro that seem to favor lysogeny. In contrast, absent or high concentrations of (p)ppGpp favor lysis. Modest ppGpp levels favor lysogeny by leading to low HflB (FtsH). When ppGpp is either absent or high, HflB protease levels are high; this leads to lower CII (a lysogeny-promoting phage protein) and favors lysis.
Impact on transcription
Characteristics of affected promoters
One of the key elements of promoters inhibited by (p)ppGpp is the presence of a GC-rich discriminator, defined as a region between TATA-box (-10 box) and +1 nt (where +1 is the transcription start sit). Promoters negatively regulated by ppGpp have a 16-bp linker, in contrast with the 17-bp consensus. Promoters activated by ppGpp seem to have a AT-rich discriminator and linger linkers (for example, the his promoter linker is 18 bp).
RNAP is the target
Genetic evidence suggesting that RNAP was the target of ppGpp came from the discovery that M+ mutants (also called stringent RNAP mutants) display in vitro and in vivo mimicry of physiology and transcription regulation conferred by (p)ppGpp, even in its absence. Cross-linking ppGpp to RNAP reinforced this notion. Structural details of an association between ppGpp and RNAP came from the analysis of cocrystals that positioned ppGpp in the secondary channel of RNAP near the catalytic center.
DksA augments regulation
DksA is a 17-kDa protein, its structure is similar to GreA and GreB, which are well-characterized transcriptional elongation factors. GreA and GreB bind directly to RNAP rather than DNA and act by inserting their N-terminal coiled-coil finger domain through the RNAP secondary channel. Two conserved acidic residues at the tip of the finger domain are necessary to induce RNAP's intrinsic ability to cleave backtracked RNA. DksA also possesses two acidic residues at its finger tip, but it does not induce nucleolytic cleavage activity. Instead, these residues are proposed to stabilize ppGpp binding to RNAP by mutual coordination of an Mg2+ ion that is crucial for polymerization.
Transcription inhibition and activation
ppGpp directly inhibits transcription from ribosomal promoters. One model is ppGpp and DksA together and independently decrease the stability of the open complexes formed on DNA by RNAP. Another model is the trapping mechanism. In this model, RNAP is trapped by ppGpp in closed complexes and is unable to initiate transcription. Thus, ppGpp seems to act at many levels, and the mechanism of its action is a complex outcome of several factors, intrinsic promoter properties not being the least of them. The transcription activation by ppGpp can be direct or indirect. Direct activation occurs when RNAP interacts with effectors, such as ppGpp, DksA or both, to increase transcription from a given promoter. Indirect activation by these effectors of one promoter relies on inhibition of other (strong) promoters, leading to increased availability of RNAP that indirectly activates transcription initiation. The promoters that activated directly by ppGpp include PargI, PthrABC, PlivJ, and PhisG. The indirectly activation promoters include these dependent on sigma factors: S, H, N, E. When strong promoters, such as rrn, are inhibited, there more RNAP are available for these alternative sigma factors.
Pathogenesis and (p)ppGpp
When (p)ppGpp is absent, pathogenicity is compromised for reasons that vary with the organism studied. Deleting relA and spoT genes, but not relA alone, gave a (p)ppGpp0 state that resulted in strong attenuation in mice and noninvasiveness in vitro. Vaccine tests reveal that 30 days after single immunization with the (p)ppGpp0 strain, mice were protected from challenge with wild-type Salmonella at a dose 106-fold above the established LD50.
References
- ↑ Srivatsan, A.; Wang, J. D. (2008). "Control of bacterial transcription, translation and replication by (p)ppGpp". Current Opinion in Microbiology. 11 (2): 100–105. doi:10.1016/j.mib.2008.02.001. PMID 18359660.
- ↑ Cashel M, Gentry DR, Hernandez VH, Vinella D: The stringent response. In Escherichia coli and Salmonella: cellular and molecular biology, edn 2. Edited by Neidhardt FC, Curtiss III R, Ingraham JL, Lin ECC, Low KB, Magasanik B, Reznikoff WS, Riley M, Schaechter M, Umbarger HE.ASM Press; 1996.
Further reading
- Condon, C; Squires, C; Squires, CL (1995). "Control of rRNA transcription in Escherichia coli". Microbiol Rev. 59 (4): 623–45. PMC 239391
 . PMID 8531889.
. PMID 8531889. - Artsimovitch, I; Patlan, V; Sekine, S; Vassylyeva, MN; Hosaka, T; Ochi, K; Yokoyama, S; Vassylyev, DG (2004). "Structural basis for transcription regulation by alarmone ppGpp". Cell. 117 (3): 299–310. doi:10.1016/S0092-8674(04)00401-5. PMID 15109491.
- Magnusson, LU; Farewell, A; Nyström, T (2005). "PpGpp: A global regulator in Escherichia coli". Trends in Microbiology. 13 (5): 236–42. doi:10.1016/j.tim.2005.03.008. PMID 15866041.
- Potrykus, K; Cashel, M (2008). "(p)ppGpp: Still magical?". Annu. Rev. Microbiol. 62: 35–51. doi:10.1146/annurev.micro.62.081307.162903. PMID 18454629.