310 helix
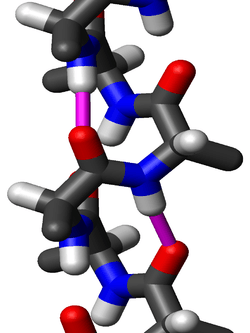
A 310 helix is a type of secondary structure found in proteins and polypeptides.Of the countless protein secondary structures present, the 310-helix is the fourth most common type observed; following α-helices, β-sheets and reverse turns. 310-helices constitute nearly 10-15% of all helices in protein secondary structures, and are typically observed as extensions of α-helices found at either their N- or C- termini. Because of the α-helices tendency to consistently fold and unfold, it has been proposed that the 310-helix serves as an intermediary conformation of sorts, and provides insight into the initiation of α-helix folding.

Discovery
Right around the 1950s, Max Perutz, who was the head of the Medical Research Council at the University of Cambridge, first wrote a paper documenting the elusive 310-helix. Of those cited in his work, two of his most famous colleagues during these experiments were James Watson and Francis Crick. In his paper, his two main observations regarding the theoretical configurations of polypeptides were as follows:
- The chains are held together by hydrogen bonding between –NH and –CO groups of nearby amino acid residues.
- Those structures in which all NH and CO groups are hydrogen bonded are inherently more probable, because their free energy is presumably lower. His analysis of these secondary structural cues were all done through collection of noncrystalline diffraction data, as well as from small molecule crystal structures such as collagen found in hair.
Structure
The amino acids in a 310-helix are arranged in a right-handed helical structure. Each amino acid corresponds to a 120° turn in the helix (i.e., the helix has three residues per turn), and a translation of 2.0 Å (= 0.2 nm) along the helical axis, and has 10 atoms in the ring formed by making the hydrogen bond. Most importantly, the N-H group of an amino acid forms a hydrogen bond with the C = O group of the amino acid three residues earlier; this repeated i + 3 → i hydrogen bonding defines a 310-helix. Similar structures include the α-helix (i + 4 → i hydrogen bonding) and the π-helix i + 5 → i hydrogen bonding.
Residues in long 310-helices adopt (φ, ψ) dihedral angles near (−49°, −26°). Many 310-helices in proteins are short, so deviate from these values. More generally, residues in long 310-helices adopt dihedral angles such that the ψ dihedral angle of one residue and the φ dihedral angle of the next residue sum to roughly −75°. For comparison, the sum of the dihedral angles for an α-helix is roughly −105°, whereas that for a π-helix is roughly −125°.
The general formula for the rotation angle Ω per residue of any polypeptide helix with trans isomers is given by the equation
- The (−49°, −26°) (φ, ψ) dihedral angles could be attributed to the short lengths of these helices, ranging anywhere from 3-5 residues long, compared to the 10-12 residue lengths of their α-helix contemporaries. Short residue lengths can cause deviations in their main-chain torsion angle distributions, accounting for their irregularity. Scientists’ theory behind the instability of 310-helices arises from their small lengths, as well as the fact that they constitute a distorted hydrogen bond network, though the frequency of the 310-helix in nature suggests that it serves an ample purpose.

Stability
Through research carried out by Mary Karpen, Pieter De Haseth and Kenneth Neet, new findings in the partial stability in 310-helices were uncovered. The helices are most noticeably stabilized by an aspartate residue at the nonpolar N-terminal that interacts with the amide group at the helical N-cap. This electrostatic interaction stabilizes the peptide dipoles in a parallel orientation. Much like the contiguous helical hydrogen bonds that stabilize α-helices, high levels of aspartate are just as equally important in the survival of the 310-helix. High frequency of aspartate in both 310-helix and α-helices is indicative of its helix initiation, but at the same time suggests that it favors stabilization of the 310-helix by inhibiting the propagation of α-helices.
See also
References
- Pauling L, Corey RB and Branson HR. (1951) "The Structure of Proteins: Two Hydrogen-Bonded Helical Configurations of the Polypeptide Chain", Proc. Natl. Acad. Sci. Wash., 37, 205.
- Toniolo C, Benedetti E. (1991) "The polypeptide 3/10-helix", Trends Biochem. Sci., 16, 350-353.
- A 310 Helix Is a Type of Protein Secondary." Biochemistries. N.p., 20 Oct. 2013. Web. 06 Dec. 2015. <http://biochemistri.es/the-3-10-helix>.
- Karpen, Mary E., Pieter L. De Haseth, and Kenneth E. Neet. "Differences in the Amino Acid Distributions of 3 10 -helices and α -helices." Protein Sci. Protein Science 1.10 (1992): 1333-342. Wiley. Web. 04 Dec. 2015.
- Vieira-Pires, R. S., and J. H. Morais-Cabral. "310 Helices in Channels and Other Membrane Proteins." The Journal of General Physiology 136.6 (2010): 585-92. Web.
- Armen, Roger, Darwin O.v. Alonso, and Valerie Daggett. "The Role of α-, 3 10 -, and π-helix in Helix→coil Transitions." Protein Sci. Protein Science 12.6 (2003): 1145-157. Web.
| ||||||||||||||||||||||||||||||||||