Hadrosauroidea
| Hadrosauroids Temporal range: Early-Late Cretaceous, 130–66 Ma | |
|---|---|
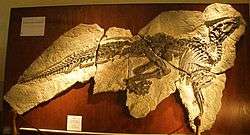 | |
| Holotype skeleton of Tethyshadros insularis | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Dinosauria |
| Order: | †Ornithischia |
| Suborder: | †Ornithopoda |
| Clade: | †Hadrosauriformes |
| Superfamily: | †Hadrosauroidea Cope, 1869 |
| Type species | |
| †Hadrosaurus foulkii Leidy, 1858 | |
| Families | |
|
†Hadrosauridae | |
| Synonyms | |
Hadrosauroidea is a clade or superfamily of ornithischian dinosaurs that includes the "duck-billed" dinosaurs, or hadrosaurids, and all dinosaurs more closely related to them than to Iguanodon. Many primitive hadrosauroids, such as the sail-backed Ouranosaurus, have traditionally been included in a paraphyletic (unnatural grouping) "Iguanodontidae". With cladistic analysis, the traditional Iguanodontidae has been largely disbanded, and probably includes only Iguanodon and perhaps its closest relatives.
Classification
Taxonomy
The following taxonomy follows Ramírez-Velasco et al, 2012 unless otherwise noted.[1]
- Superfamily Hadrosauroidea
- Altirhinus[2]
- Bactrosaurus
- Batyrosaurus[3]
- Bolong[3]
- Claosaurus
- Equijubus
- Eolambia
- Fukuisaurus
- Gilmoreosaurus
- Glishades
- Huehuecanauhtlus
- Jeyawati
- Jintasaurus
- Jinzhousaurus
- Koshisaurus[4]
- Levnesovia
- Lophorhothon
- Nanyangosaurus
- Penelopognathus
- Probactrosaurus
- Protohadros
- Siamodon
- Shuangmiaosaurus
- Tanius
- Tethyshadros
- Xuwulong[5]
- Family Hadrosauridae
Phylogeny
The cladogram below follows an analysis by Wu Wenhao and Pascal Godefroit (2012).[3]
| Hadrosauriformes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Cladogram after Prieto-Marquez and Norell (2010).[6]
| Hadrosauroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
A phylogenetic analysis performed by Ramírez-Velasco et al. (2012) found a big polytomy of all hadrosauroids which are more derived than Probactrosaurus but less derived than Hadrosauridae. The exclusion of Claosaurus, Jeyawati, Levnesovia, Nanyangosaurus, Shuangmiaosaurus and Telmatosaurus from the polytomy resulted in a more resolved topology. The cladogram below shows Huehuecanauhtlus phylogenetic position among other hadrosauroids following this topology (the relationships within Hadrosauridae are not shown).[1]
| Hadrosauroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
See also
References
- 1 2 Angel Alejandro Ramírez-Velasco; Mouloud Benammi; Albert Prieto-Márquez; Jesús Alvarado Ortega; René Hernández-Rivera (2012). "Huehuecanauhtlus tiquichensis, a new hadrosauroid dinosaur (Ornithischia: Ornithopoda) from the Santonian (Late Cretaceous) of Michoacán, Mexico". Canadian Journal of Earth Sciences. 49 (2): 379–395. doi:10.1139/e11-062.
- ↑ Hai-Lu You; Da-Qing Li (2009). "A new basal hadrosauriform dinosaur (Ornithischia: Iguanodontia) from the Early Cretaceous of northwestern China". Canadian Journal of Earth Sciences. 46 (12): 949–957. doi:10.1139/E09-067.
- 1 2 3 Wu Wenhao; Pascal Godefroit (2012). "Anatomy and Relationships of Bolong yixianensis, an Early Cretaceous Iguanodontoid Dinosaur from Western Liaoning, China". In Godefroit, P. Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Indiana University Press. pp. 293–333.
- ↑ Shibaba, Masateru; Azuma, Yoichi (2015). "New basal hadrosauroid (Dinosauria: Ornithopoda) from the Lower Cretaceous Kitadani Formation, Fukui, central Japan." (PDF). Zootaxa. 3914: 421. doi:10.11646/zootaxa.3914.4.3.|
- ↑ McDonald, A. T. (2012). Farke, Andrew A, ed. "Phylogeny of Basal Iguanodonts (Dinosauria: Ornithischia): An Update". PLoS ONE. 7 (5): e36745. doi:10.1371/journal.pone.0036745. PMC 3358318
 . PMID 22629328.
. PMID 22629328. - ↑ Albert Prieto-Marquez and Mark A. Norell (2010). "Anatomy and Relationships of Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) from the Late Cretaceous of Central Asia" (PDF). American Museum Novitates. 3694: 1–52. doi:10.1206/3694.2. ISSN 0003-0082.