Rate of evolution
The rate of evolution is a variable of considerable interest in evolutionary biology. It concerns the limits of adaptation to natural environments as well as the limits of artificial selection.







Humans have created a wide range of new species, and varieties within those species, of both domesticated animals and plants. Other human activity also impacts evolution.[1] This has been achieved in a very short geological period of time, spanning only a few tens of thousands of years, and sometimes less. Maize, Zea mays, for instance, is estimated to have been created in what is now known as Mexico in only a few thousand years, starting between about 7 000 and 12 000 years ago, from still uncertain origins.[2] In the light of this extraordinarily rapid rate of evolution, through (prehistoric) artificial selection, George C. Williams[3] and others,[4][5][6] have remarked the following:
The question of evolutionary change in relation to available geological time is indeed a serious theoretical challenge, but the reasons are exactly the opposite of that inspired by most people’s intuition. Organisms in general have not done nearly as much evolving as we should reasonably expect. Long term rates of change, even in lineages of unusual rapid evolution, are almost always far slower than they theoretically could be. The basis for such expectation is to be found most clearly in observed rates of evolution under artificial selection, along with the often high rates of change in environmental conditions that must imply rapid change in intensity and direction of selection in nature.[3]
Evolution is imposed on species or groups. It is not planned or striven for in some Lamarckist way.[7] The mutations on which the process depends are random events, and, except for the "silent mutations" which do not affect the functionality or appearance of the carrier, are thus usually disadvantageous, and their chance of proving to be useful in the future is vanishingly small. Therefore, while a species or group might benefit from being able to adapt to a new environment by accumulating a wide range of genetic variation, this is to the detriment of the individuals who have to carry these mutations until a small, unpredictable minority of them ultimately contributes to such an adaptation. Thus, the capability to evolve is a group adaptation, a concept discredited by (for example) George C. Williams,[8] John Maynard Smith[9] and Richard Dawkins[10][11][12][13] as selectively disadvantageous to the individual. .
If sexual creatures avoid mates with strange or unusual characteristics, in the process called koinophilia,[14][15][16][17] then mutations that affect the external appearance of their carriers will seldom be passed on to the next and subsequent generations. They will therefore seldom be tested by natural selection. Evolution is, therefore, effectively halted or slowed down considerably. The only mutations that can accumulate in a population are ones that have no noticeable effect on the outward appearance and functionality of their bearers (i.e., they are "silent" or "neutral mutations", which can be, and are, used to trace the relatedness and age of populations and species.[14][18])
This implies that evolution can only occur if mutant mates cannot be avoided, as a result of a severe scarcity of potential mates. This is most likely to occur in small, isolated communities. These occur most commonly on small islands, in remote valleys, lakes, river systems, or caves,[19] or during the aftermath of a mass extinction.[18] Under these circumstances, not only is the choice of mates severely restricted but population bottlenecks, founder effects, genetic drift and inbreeding cause rapid, random changes in the isolated population's genetic composition.[19] Furthermore, hybridization with a related species trapped in the same isolate might introduce additional genetic changes. If an isolated population such as this survives its genetic upheavals, and subsequently expands into an unoccupied niche, or into a niche in which it has an advantage over its competitors, a new species, or subspecies, will have come in being. In geological terms this will be an abrupt event. A resumption of avoiding mutant mates will, thereafter, result, once again, in evolutionary stagnation.
Thus the fossil record of an evolutionary progression typically consists of species that suddenly appear, and ultimately disappear, in many cases close to a million years later, without any change in external appearance.[18][20][21] Graphically, these fossil species are represented by horizontal lines, whose lengths depict how long each of them existed. The horizontality of the lines illustrates the unchanging appearance of each of the fossil species depicted on the graph. During each species' existence new species appear at random intervals, each also lasting many hundreds of thousands of years before disappearing without a change in appearance. The exact relatedness of these concurrent species is generally impossible to determine. This is illustrated in the following diagram depicting the evolution of modern humans from the time that the Hominins separated from the line that led to the evolution of our closest living primate relatives, the chimpanzees.[21]
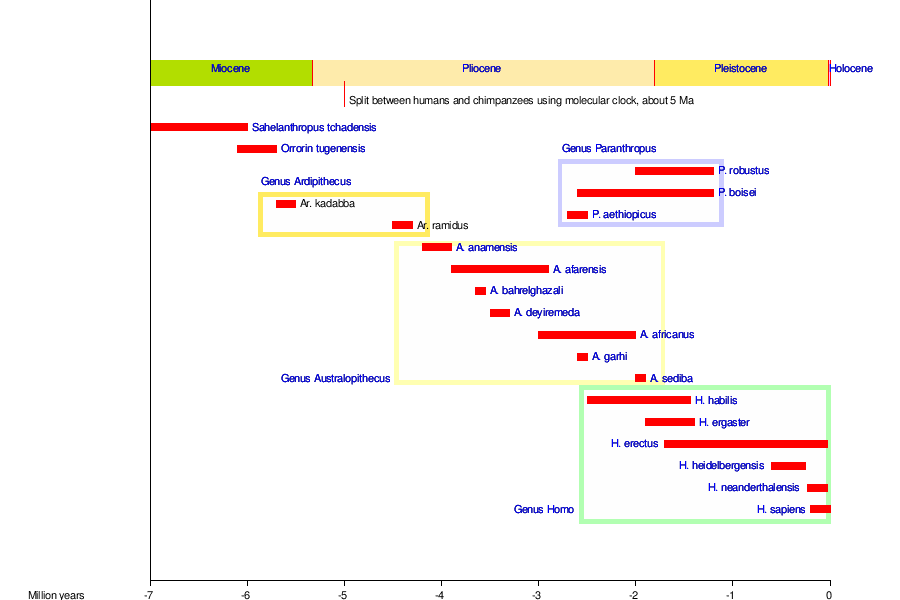 |
For similar evolutionary time lines see, for instance, the paleontological list of African dinosaurs, Asian dinosaurs, the Lampriformes and Amiiformes.
See also
References
- ↑ Milius, Susan (June 24, 2016). "Cities create accidental experiments in plant, animal evolution; From clover to lizards, a city's nonhuman residents evolve in human-scale time". Science News. Retrieved July 25, 2016.
- ↑ Laws, B (2010).Fifty Plants that Changed the Course of History. pp. 210- 215. Cinincinnati, David and Charles Book
- 1 2 Williams, G.C. (1992). Stasis. In Natural Selection: Domains, Levels and Challenges. p. 128. New York: Oxford University Press.
- ↑ Eldredge, N., Gould, S.J. (1972). "Punctuated equilibria: an alternative to phyletic gradualism" In Schopf, T.J.M., ed., Models in Paleobiology. San Francisco: Freeman Cooper. pp. 82–115. Reprinted in Eldredge, N. (1985) Time frames. Princeton: Princeton Univ. Press.
- ↑ Mayr, E. (1954). Changes of genetic environment and evolution. In: Evolution as a process Eds. Huxley, J., Hardy, A.C., Ford, E.B. pp. 157-180. New York: Columbia University Press.
- ↑ Maynard Smith, J. (1989). Evolutionary Genetics p. 281. Oxford: Oxford University Press.
- ↑ Gould, S.J. (1980) Return of the Hopeful Monster. in The Panda's Thumb. p. 186-193. New York: W.W. Norton.
- ↑ Williams, G.C. (1972) Adaptation and Natural Selection: A Critique of Some Current Evolutionary Thought. Princeton University Press.ISBN 0-691-02357-3
- ↑ Maynard Smith, J. (1964). "Group selection and kin selection". Nature. 201 (4924): 1145–1147. doi:10.1038/2011145a0.
- ↑ See the chapter God's utility function in Dawkins, Richard (1995). River Out of Eden. New York: Basic Books. ISBN 0-465-06990-8.
- ↑ Dawkins, R. (1994). "Burying the Vehicle Commentary on Wilson & Sober: Group Selection". Behavioural and Brain Sciences. 17 (4): 616–617. Archived from the original on 2006-09-15.
- ↑ Dennett, D.C. (1994). "E Pluribus Unum? Commentary on Wilson & Sober: Group Selection". Behavioural and Brain Sciences. 17 (4): 617–618. Archived from the original on 2007-12-27.
- ↑ Pinker, S. (2012). The False Allure of Group Selection. Edge, Jun 19, 2012. http://edge.org/conversation/the-false-allure-of-group-selection
- 1 2 Koeslag, J.H. (1990). "Koinophilia groups sexual creatures into species, promotes stasis, and stabilizes social behaviour." J. theor. Biol. 144, 15–35
- ↑ Miller, W.B. (2013). "What is the big deal about evolutionary gaps?". In: The Microcosm within: Evolution and Extinction in the Hologenome. Boca Raton, Florida.: Universal Publishers. pp. 177, 395–396. ISBN 1-61233-2773.
- ↑ Symons, D. (1979) The Evolution of Human Sexuality. Oxford: Oxford University Press.
- ↑ Langlois, J.H., Roggman, L. (1990). "Attractive faces are only average." Psychol. Sci. 1, 115–121
- 1 2 3 Campbell, N.A. (1990) Biology p. 450–451, 487–490, 499–501. Redwood City CA: Benjamin Cummings Publishing Company.
- 1 2 Ayala, F.J. (1982) Population and Evolutionary Genetics pp. 73–83, 182–190, 198–215. Menlo Park, California: Benjamin/Cummings. ISBN 0-8053-0315-4
- ↑ Gould, S.J. & Eldredge, N. (1977) "Punctuated equilibria: the tempo and mode of evolution reconsidered." Paleobiology 3 115–151.
- 1 2 McCarthy, T. & Rubridge, B. (2005) The Story of Earth and Life. Cape Town: Struik Publishers. ISBN 1-77007-148-2.