Archosaur
| Archosaurs Temporal range: Early Triassic–Present, 250–0 Ma | |
|---|---|
 | |
| Birds and crocodilians (in this case a yellow-billed stork and a Nile crocodile) are the only living archosaur groups. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Eucrocopoda |
| Clade: | Archosauria Cope, 1869 |
| Subgroups | |
| |
| Synonyms | |
|
Arctopoda Haeckel, 1895 | |
Archosaurs are a group of diapsid amniotes whose living representatives consist of birds and crocodilians. This group also includes all extinct non-avian dinosaurs, extinct crocodilian relatives, and pterosaurs. Archosauria, the archosaur clade, is a crown group that includes the most recent common ancestor of living birds and crocodilians. It includes two main clades: Pseudosuchia, which includes crocodilians and their extinct relatives, and Ornithosuchia, which includes birds and their extinct relatives (such as non-avian dinosaurs and pterosaurs).
Distinguishing characteristics
Archosaurs can be distinguished from other tetrapods on the basis of several synapomorphies, or shared characteristics, first found in a common ancestor. The simplest and most widely agreed synapomorphies of archosaurs include teeth set in sockets, antorbital and mandibular fenestrae (openings in front of the eyes and in the jaw, respectively),[1] and a fourth trochanter (a prominent ridge on the femur).[2] Being set in sockets, the teeth were less likely to be torn loose during feeding. This feature is responsible for the name "thecodont" (meaning "socket teeth"),[3] which paleontologists used to apply to many Triassic archosaurs.[2] Some archosaurs, such as birds, are secondarily toothless. Antorbital fenestrae reduced the weight of the skull, which was relatively large in early archosaurs, rather like that of modern crocodilians. Mandibular fenestrae may also have reduced the weight of the jaw in some forms. The fourth trochanter provides a large site for the attachment of muscles on the femur. Stronger muscles allowed for erect gaits in early archosaurs, and may also be connected with the ability of the archosaurs or their immediate ancestors to survive the catastrophic Permian-Triassic extinction event.
Origins
There is some debate about when archosaurs first appeared. Those who classify the Permian reptiles Archosaurus rossicus and/or Protorosaurus speneri as true archosaurs maintain that archosaurs first appeared in the late Permian. Some taxonomists classify both Archosaurus rossicus and Protorosaurus speneri as archosauriforms (not true archosaurs but very closely related); these taxonomists maintain that archosaurs first evolved from archosauriform ancestors during the Olenekian stage of the Early Triassic. The earliest archosaurs were rauisuchians, such as Scythosuchus and Tsylmosuchus, both of which have been found from Russia and date back to the Olenekian.[4]
Archosaur takeover in the Triassic

Synapsids (a group including mammals and their extinct relatives, which are often referred to as "mammal-like reptiles") were the dominant land vertebrates throughout the Permian, but most perished in the Permian-Triassic extinction event. Very few large synapsids survived the event, although one form, Lystrosaurus (an herbivorous dicynodont), attained a widespread distribution soon after the extinction.
But archosaurs quickly became the dominant land vertebrates in the early Triassic. The two most commonly suggested explanations for this are:
- Archosaurs made more rapid progress towards erect limbs than synapsids, and this gave them greater stamina by avoiding Carrier's constraint. An objection to this explanation is that archosaurs became dominant while they still had sprawling or semi-erect limbs, similar to those of Lystrosaurus and other synapsids.
- The early Triassic was predominantly arid, because most of the earth's land was concentrated in the supercontinent Pangaea. Archosaurs were probably better at conserving water than early synapsids because:
- Modern diapsids (lizards, snakes, crocodilians, birds) excrete uric acid, which can be excreted as a paste. It is reasonable to suppose that archosaurs (the ancestors of crocodilians, dinosaurs and birds) also excreted uric acid, and therefore were good at conserving water. The aglandular (glandless) skins of diapsids would also have helped to conserve water.
- Modern mammals excrete urea, which requires a lot of water to keep it dissolved. Their skins also contain many glands, which also lose water. Assuming that early synapsids had similar features, e.g., as argued by the authors of Palaeos, they were at a disadvantage in a mainly arid world. The same well-respected site points out that "for much of Australia's Plio-Pleistocene history, where conditions were probably similar, the largest terrestrial predators were not mammals but gigantic varanid lizards (Megalania) and land crocs."[3]
However, this theory has been questioned, since it implies synapsids were necessarily less advantaged in water retention, that synapsid decline coincides with climate changes or archosaur diversity (neither of which tested) and the fact that desert dwelling mammals as well adapted in this department as archosaurs,[5] and some cynodonts like Trucidocynodon were large sized predators.[6]
Main forms
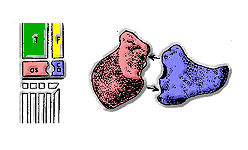



Since the 1970s, scientists have classified archosaurs mainly on the basis of their ankles.[7] The earliest archosaurs had "primitive mesotarsal" ankles: the astragalus and calcaneum were fixed to the tibia and fibula by sutures and the joint bent about the contact between these bones and the foot.

The Pseudosuchia appeared early in the Triassic. In their ankles, the astragalus was joined to the tibia by a suture and the joint rotated round a peg on the astragalus which fitted into a socket in the calcaneum. Early "crurotarsans" still walked with sprawling limbs, but some later crurotarsans developed fully erect limbs (most notably the Rauisuchia). Modern crocodilians are crurotarsans that can walk with their limbs sprawling or erect depending on speed of locomotion.
Euparkeria and the Ornithosuchidae had "reversed crurotarsal" ankles, with a peg on the calcaneum and socket on the astragalus.

The earliest fossils of Avemetatarsalia ("bird ankles") appear in the Carnian age of the late Triassic, but it is hard to see how they could have evolved from crurotarsans — possibly they actually evolved much earlier, or perhaps they evolved from the last of the "primitive mesotarsal" archosaurs. Ornithodires' "advanced mesotarsal" ankle had a very large astragalus and very small calcaneum, and could only move in one plane, like a simple hinge. This arrangement, which was only suitable for animals with erect limbs, provided more stability when the animals were running. The ornithodires differed from other archosaurs in other ways: they were lightly built and usually small, their necks were long and had an S-shaped curve, their skulls were much more lightly built, and many ornithodires were completely bipedal. The archosaurian fourth trochanter on the femur may have made it easier for ornithodires to become bipeds, because it provided more leverage for the thigh muscles. In the late Triassic, the ornithodires diversified to produce dinosaurs and pterosaurs.
Classification
Modern classification
Archosauria is normally defined as a crown group, which means that it only includes descendants of the last common ancestors of its living representatives. In the case of archosaurs, these are birds and crocodilians. Archosauria is within the larger clade Archosauriformes, which includes some close relatives of archosaurs, such as proterochampsids and euparkeriids. These relatives are often referred to as archosaurs despite being placed outside of the crown group Archosauria in a more basal position within Archosauriformes.[8] Historically, many archosauriforms were described as archosaurs, including proterosuchids and erythrosuchids, based on the presence of an antorbital fenestra. While many researchers prefer to treat Archosauria as an unranked clade, some continue to assign it a traditional biological rank. Traditionally, Archosauria has been treated as a Superorder, though a few 21st century researchers have assigned it to different ranks including Division[9] and Class.[10]
History of classification
Archosauria as a term was first coined by American paleontologist Edward Drinker Cope in 1869, and included a wide range of taxa including dinosaurs, crocodilians, thecodonts, sauropterygians (which may be related to turtles), rhynchocephalians (a group that according to Cope included rhynchosaurs, which nowadays are considered to be more basal archosauromorphs, and tuataras, which are lepidosaurs), and anomodonts, which are now considered synapsids.[11] It was not until 1986 that Archosauria was defined as a crown-clade, restricting its use to more derived taxa.[12]
Cope's term was a Greek-Latin hybrid intended to refer to the cranial arches, but has later also been understood as "leading reptiles" or "ruling reptiles" by association with Greek ἀρχός "leader, ruler".[13]
The term "thecodont", now considered an obsolete term, was first used by the English paleontologist Richard Owen in 1859 to describe Triassic archosaurs, and it became widely used in the 20th century. Thecodonts were considered the "basal stock" from which the more advanced archosaurs descended. They did not possess features seen in later avian and crocodilian lines, and therefore were considered more primitive and ancestral to the two groups. With the cladistic revolution of the 1980s and 90s, in which cladistics became the most widely used method of classifying organisms, thecodonts were no longer considered a valid grouping. Because they are considered a "basal stock", thecodonts are paraphyletic, meaning that they form a group that does not include all descendants of its last common ancestor: in this case, the more derived crocodilians and birds are excluded from "Thecodontia" as it was formerly understood. The description of the basal ornithodires Lagerpeton and Lagosuchus in the 1970s provided evidence that linked thecodonts with dinosaurs, and contributed to the disuse of the term "Thecodontia", which many cladists consider an artificial grouping.[14]
With the identification of "crocodilian normal" and "crocodilian reversed" ankles by Sankar Chatterjee in 1978, a basal split in Archosauria was identified. Chatterjee considered these two groups to be Pseudosuchia with the "normal" ankle and Ornithosuchidae with the "reversed" ankle. Ornithosuchids were thought to be ancestral to dinosaurs at this time. In 1979, A.R.I. Cruickshank identified the basal split and thought that the crurotarsan ankle developed independently in these two groups, but in opposite ways. Cruickshank also thought that the development of these ankle types progressed in each group to allow advanced members to have semi-erect (in the case of crocodilians) or erect (in the case of dinosaurs) gaits.[14]
Phylogeny
In many phylogenetic analyses, archosaurs have been shown to be a monophyletic grouping, thus forming a true clade. One of the first studies of archosaur phylogeny was authored by French paleontologist Jacques Gauthier in 1986. Gauthier split Archosauria into Pseudosuchia, the crocodilian line, and Ornithosuchia, the dinosaur and pterosaur line. Pseudosuchia was defined as all archosaurs more closely related to crocodiles, while Ornithosuchia was defined as all archosaurs more closely related to birds. Proterochampsids, erythrosuchids, and proterosuchids fell successively outside Archosauria in the resulting tree. Below is the cladogram from Gauthier (1986):[15]
| |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
In 1988, paleontologists Michael Benton and J.M. Clark produced a new tree in a phylogenetic study of basal archosaurs. As in Gauthier's tree, Benton and Clark's revealed a basal split within Archosauria. They referred to the two groups as Crocodylotarsi and Ornithosuchia. Crocodylotarsi was defined as an apomorphy-based taxon based on the presence of a "crocodile-normal" ankle joint (considered to be the defining apomorphy of the clade). Gauthier's Pseudosuchia, by contrast, was a stem-based taxon. Unlike Gauthier's tree, Benton and Clark's places Euparkeria outside Ornithosuchia and outside the crown group Archosauria altogether.[16]
The clades Crurotarsi and Ornithodira were first used together in 1990 by paleontologist Paul Sereno and A.B. Arcucci in their phylogenetic study of archosaurs. They were the first to erect the clade Crurotarsi, while Ornithodira was named by Gauthier in 1986. Crurotarsi and Ornithodira replaced Pseudosuchia and Ornithosuchia, respectively, as the monophyly of both of these clades were questioned.[14][17] Sereno and Arcucci incorporated archosaur features other than ankle types in their analyses, which resulted in a different tree than previous analyses. Below is a cladogram based on Sereno (1991), which is similar to the one produced by Sereno and Arcucci:[14]
| Archosauriformes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Ornithodira and Crurotarsi are both node-based clades, meaning that they are defined to include the last common ancestor of two or more taxa and all of its descendants. Ornithodira includes the last common ancestor of pterosaurs and dinosaurs (which include birds), while Crurotarsi includes the last common ancestor of living crocodilians and three groups of Triassic archosaurs: ornithosuchids, aetosaurs, and phytosaurs. These clades are not equivalent to "bird-line" and "crocodile-line" archosaurs, which would be branch-based clades defined as all taxa more closely related to one living group (either birds or crocodiles) than the another.

Benton proposed the name Avemetatarsalia in 1999 to include all bird-line archosaurs (under his definition, all archosaurs more closely related to dinosaurs than to crocodilians). His analysis of the small Triassic archosaur Scleromochlus placed it within bird-line archosaurs but outside Ornithodira, meaning that Ornithodira was no longer equivalent to bird-line archosaurs. Below is a cladogram modified from Benton (2004) showing this phylogeny:[12]
| Archosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
In Sterling Nesbitt's 2011 monograph on early archosaurs, a phylogenetic analysis found strong support for phytosaurs falling outside Archosauria. Many subsequent studies supported this phylogeny. Because Crurotarsi is defined by the inclusion of phytosaurs, the placement of phytosaurs outside Archosauria means that Crurotarsi must include all of Archosauria. Nesbitt reinstated Pseudosuchia as a clade name for crocodile-line archosaurs, using it as a stem-based taxon. Below is a cladogram modified from Nesbitt (2011):[18]
| |
* Nesbitt did not include Scleromochlus in the analysis, meaning that Avemetatarsalia and Ornithodira occupy the same place in this cladogram | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Extinction and survival
Crocodilians, pterosaurs and dinosaurs survived the Triassic–Jurassic extinction event about 200 million years ago, but other archosaurs became extinct.
Non-avian dinosaurs and pterosaurs perished in the Cretaceous–Paleogene extinction event, which occurred approximately 66 million years ago, but birds (the only remaining dinosaur group) and crocodilians survived. Both are descendants of archosaurs, and are therefore archosaurs themselves under phylogenetic taxonomy.
Crocodilians (which include all modern crocodiles, alligators, and gharials) and birds flourish today. It is generally agreed that birds have the most species of all terrestrial vertebrates.
Archosaur lifestyle
Hip joints and locomotion
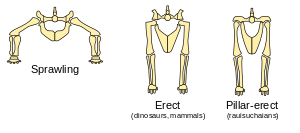
Like the early tetrapods, early archosaurs had a sprawling gait because their hip sockets faced sideways, and the knobs at the tops of their femurs were in line with the femur.
In the early to middle Triassic, some archosaur groups developed hip joints that allowed (or required) a more erect gait. This gave them greater stamina, because it avoided Carrier's constraint, i.e. they could run and breathe easily at the same time. There were two main types of joint which allowed erect legs:
- The hip sockets faced sideways, but the knobs on the femurs were at right angles to the rest of the femur, which therefore pointed downwards. Dinosaurs evolved from archosaurs with this hip arrangement.
- The hip sockets faced downwards and the knobs on the femurs were in line with the femur. This "pillar-erect" arrangement appears to have evolved independently in various archosaur lineages, for example it was common in Rauisuchia and also appeared in some aetosaurs.
It has been pointed out that an upright stance requires more energy, so it may indicate a higher metabolism and a higher body temperature.[19]
Diet
Most were large predators, but members of various lines diversified into other niches. Aetosaurs were herbivores and some developed extensive armor. A few crocodilians were herbivores, e.g., Simosuchus, Phyllodontosuchus. The large crocodilian Stomatosuchus may have been a filter feeder. Sauropodomorphs and ornithischian dinosaurs were herbivores with diverse adaptations for feeding biomechanics.
Land, water and air
Archosaurs are mainly portrayed as land animals, but:
- The phytosaurs and crocodilians dominated the rivers and swamps and even invaded the seas (e.g., the teleosaurs, Metriorhynchidae and Dyrosauridae). The Metriorhynchidae were rather dolphin-like, with paddle-like forelimbs, a tail fluke and smooth, unarmoured skins.
- Two clades of ornithodirans, the pterosaurs and the birds, dominated the air after becoming adapted to a volant lifestyle.
Metabolism
The metabolism of archosaurs is still a controversial topic. They certainly evolved from cold-blooded ancestors, and the surviving non-dinosaurian archosaurs, crocodilians, are cold-blooded. But crocodilians have some features which are normally associated with a warm-blooded metabolism because they improve the animal's oxygen supply:
- 4-chambered hearts. Both birds and mammals have 4-chambered hearts, which completely separate the flows of oxygenated and de-oxygenated blood. Non-crocodilian reptiles have 3-chambered hearts, which are less efficient because they let the two flows mix and thus send some de-oxygenated blood out to the body instead of to the lungs. Modern crocodilians' hearts are 4-chambered, but are smaller relative to body size and run at lower pressure than those of modern birds and mammals. They also have a pulmonary bypass, which makes them functionally 3-chambered when under water, conserving oxygen.
- a secondary palate, which allows the animal to eat and breathe at the same time.
- a hepatic piston mechanism for pumping the lungs. This is different from the lung-pumping mechanisms of mammals and birds, but similar to what some researchers claim to have found in some dinosaurs.[20][21]
So, why did natural selection favour the development of these features, which are very important for active warm-blooded creatures, but of little apparent use to cold-blooded aquatic ambush predators that spend the vast majority of their time floating in water or lying on river banks?
Paleontological evidence shows that the ancestors of living crocodilians were active and endothermic (warm-blooded). Some experts believe that their archosaur ancestors were warm-blooded as well. Physiological, anatomical, and developmental features of the crocodilian heart support the paleontological evidence and show that the lineage reverted to ectothermy when it invaded the aquatic, ambush predator niche. Crocodilian embryos develop fully 4-chambered hearts at an early stage. Modifications to the growing heart form a pulmonary bypass shunt that includes the left aortic arch, which originates from the right ventricle, the foramen of Panizza between the left and right aortic arches, and the cog‐tooth valve at the base of the pulmonary artery. The shunt is used during diving to make the heart function as 3-chambered heart, providing the crocodilian with the neurally controlled shunting used by ectotherms. The researchers concluded that the ancestors of living crocodilians had fully 4-chambered hearts, and were therefore warm-blooded, before they reverted to a cold-blooded or ectothermic metabolism. The authors also provide other evidence for endothermy in stem archosaurs.[22][23] It is reasonable to suggest that later crocodilians developed the pulmonary bypass shunt as they became cold-blooded, aquatic, and less active.
If the original crocodilians and other Triassic archosaurs were warm-blooded, this would help to resolve some evolutionary puzzles:
- The earliest crocodilians, e.g., Terrestrisuchus, were slim, leggy terrestrial predators whose build suggests a fairly active lifestyle, which requires a fairly fast metabolism. And some other crurotarsan archosaurs appear to have had erect limbs, while those of rauisuchians are very poorly adapted for any other posture. Erect limbs are advantageous for active animals because they avoid Carrier's constraint, but disadvantageous for more sluggish animals because they increase the energy costs of standing up and lying down.
- If early archosaurs were completely cold-blooded and (as seems most likely) dinosaurs were at least fairly warm-blooded, dinosaurs would have had to evolve warm-blooded metabolisms in less than half the time it took for synapsids to do the same.
Respiratory system
A recent study of the lungs of the American alligator has shown that the airflow through them is unidirectional, moving in the same direction during inhalation and exhalation.[24] This is also seen in birds and many non-avian dinosaurs, which have air sacs to further aid in respiration. Both birds and alligators achieve unidirectional air flow through the presence of parabronchi, which are responsible for gas exchange. The study has found that in alligators, air enters through the second bronchial branch, moves through the parabronchi, and exits through the first bronchial branch. Unidirectional airflow in both birds and alligators suggests that this type of respiration was present in basal Triassic archosaurs and their non-dinosaurian descendants, including phytosaurs, aetosaurs, rauisuchians, crocodylomorphs, and pterosaurs.[24] The use of unidirectional airflow in the lungs of archosaurs may have given the group an advantage over synapsids, which had lungs where air moved tidally in and out through a network of bronchi that terminated in alveoli, which were cul-de-sacs. The better efficiency in gas transfer seen in archosaur lungs may have been advantageous during the times of low atmospheric oxygen which are thought to have existed during the Mesozoic.[25]
References
- ↑ Gareth Dyke; Gary Kaiser, eds. (2011). Living Dinosaurs: The Evolutionary History of Modern Birds. John Wiley & Sons. p. 10. ISBN 0470656662.
- 1 2 Khanna, D.R. (2004). Biology Of Reptiles. Discovery Publishing House. pp. 78ff. ISBN 8171419070.
- 1 2 White, T.; Kazlev, M. A. "Archosauromorpha: Overview". Palaeos.com. Archived from the original on December 20, 2010. Retrieved 6 September 2012.
- ↑ Gower, D. J.; Sennikov, A. G. (2003). "Early archosaurs from Russia". In Benton, M.J.; Shishkin, M.A.; Unwin, D.M. The Age of Dinosaurs in Russia and Mongolia. Cambridge: Cambridge University Press. pp. 140–159.
- ↑ Darren Naish, Episode 38: A Not Too Shabby Podcarts
- ↑ Oliveira, T.V.; Soares, M.B.; Schultz, C.L. (2010). "Trucidocynodon riograndensis gen. nov. et sp. nov. (Eucynodontia), a new cynodont from the Brazilian Upper Triassic (Santa Maria Formation)". Zootaxa 2382: 1–71.
- ↑ Archosauromorpha: Archosauria - Palaeos
- ↑ Gower, D. J.; Wilkinson, M. (1996). "Is there any consensus on basal archosaur phylogeny?" (PDF). Proceedings of the Royal Society B. 263 (1375): 1399–1406. doi:10.1098/rspb.1996.0205.
- ↑ Benton, M.J. (2005). Vertebrate Paleontology, 3rd ed. Blackwell Science Ltd
- ↑ Göhlich, U.B., Chiappe, L.M., Clark, J.M., and Sues, H.-D. (2005). "The systematic position of the Late Jurassic alleged dinosaur Macelognathus (Crocodylomorpha: Sphenosuchia)." Canadian Journal of Earth Sciences, 42(3): 307-321.
- ↑ Cope, Edward Drinker (1869). "Synopsis of the extinct Batrachia, Reptilia and Aves of North America". Transactions of the America Philosophical Society. 14: 1–252.
- 1 2 Benton, M.J. (2004). "Origin and relationships of Dinosauria". In Weishampel, D.B.; Dodson, P.r; Osmólska, H. The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 7–19. ISBN 0-520-24209-2.
- ↑ Pamphlets on Biology: Kofoid collection, vol. 2900 (1878), p. 731
- 1 2 3 4 Sereno, P.C. (1991). "Basal archosaurs: phylogenetic relationships and functional implications". Memoir (Society of Vertebrate Paleontology). 2: 1–53. doi:10.2307/3889336. JSTOR 3889336.
- ↑ Gauthier, J.A. (1986). "Saurischian monophyly and the origin of birds". In Padian, K. The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences. 8. San Francisco: California Academy of Sciences. pp. 1–55.
- ↑ Benton, M. J.; Clark, J. M. (1985). "Archosaur phylogeny and the relationships of the Crocodylia". In Benton, M.J. The Phylogeny and Classification of the Tetrapods. 1. Oxford: Clarendon Press. pp. 295–338. ISBN 0-19-857712-5.
- ↑ Sereno, P. C.; Arcucci, A. B. (1990). "The monophyly of crurotarsal archosaurs and the origin of bird and crocodile ankle joints". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 180: 21–52.
- ↑ Nesbitt, S.J. (2011). "The early evolution of archosaurs: relationships and the origin of major clades" (PDF). Bulletin of the American Museum of Natural History. 352: 1–292. doi:10.1206/352.1.
- ↑ Desmond, Adrián J., The hot-blooded dinosaurs: a revolution in palaeontology. 1976, Dial Press, Page 87.
- ↑ Ruben, J.; et al. (1996). "The metabolic status of some Late Cretaceous dinosaurs". Science. 273 (273): 120–147. doi:10.1126/science.273.5279.1204.
- ↑ Ruben, J.; et al. (1997). "Lung structure and ventilation in theropod dinosaurs and early birds". Science. 278 (278): 1267–1247. doi:10.1126/science.278.5341.1267.
- ↑ Seymour, R. S.; Bennett-Stamper, C. L.; Johnston, S. D.; Carrier, D. R. & Grigg, G. C. (2004). "Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution". Physiol. Biochem. Zool. 77 (6): 1051–1067. doi:10.1086/422766. PMID 15674775.
- ↑ Summers, A.P. (2005). "Evolution: Warm-hearted crocs". Nature. 434 (7035): 833–834. doi:10.1038/434833a. PMID 15829945.
- 1 2 Farmer, C. G.; Sanders, K. (2010). "Unidirectional airflow in the lungs of alligators". Science. 327 (5963): 338–340. doi:10.1126/science.1180219. PMID 20075253.
- ↑ Lisa Grossman (January 14, 2010). "Alligators breathe like birds". Science News. Retrieved January 14, 2010.
Sources
- Benton, M. J. (2004). Vertebrate Paleontology (3rd ed.). Blackwell Science.
- Carroll, R. L. (1988). Vertebrate Paleontology and Evolution'. New York: W. H. Freeman.
External links
- UCMP
- Paleos reviews the messy history of archosaur phylogeny (family tree) and has an excellent image of the various archosaur ankle types.
- Mikko's Phylogeny Archive Archosauria
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||