Evolution of photosynthesis
The evolution of photosynthesis refers to the origin and subsequent evolution of photosynthesis, the process by which light energy from the sun is used to synthesize sugars from carbon dioxide, releasing oxygen as a waste product.
The first photosynthetic organisms probably evolved early in the evolutionary history of life and most likely used reducing agents such as hydrogen or hydrogen sulfide as sources of electrons, rather than water.[1] There are three major metabolic pathways by which photosynthesis is carried out: C3 photosynthesis, C4 photosynthesis, and CAM photosynthesis. C3 photosynthesis is the oldest and most common form.
Origin
The biochemical capacity to use water as the source for electrons in photosynthesis evolved once, in a common ancestor of extant cyanobacteria.[2] The geological record indicates that this transforming event took place early in Earth's history, at least 2450–2320 million years ago (Ma), and, it is speculated, much earlier.[3][4] Available evidence from geobiological studies of Archean (>2500 Ma) sedimentary rocks indicates that life existed 3500 Ma, but the question of when oxygenic photosynthesis evolved is still unanswered. A clear paleontological window on cyanobacterial evolution opened about 2000 Ma, revealing an already-diverse biota of blue-greens. Cyanobacteria remained principal primary producers throughout the Proterozoic Eon (2500–543 Ma), in part because the redox structure of the oceans favored photoautotrophs capable of nitrogen fixation. Green algae joined blue-greens as major primary producers on continental shelves near the end of the Proterozoic, but only with the Mesozoic (251–65 Ma) radiations of dinoflagellates, coccolithophorids, and diatoms did primary production in marine shelf waters take modern form. Cyanobacteria remain critical to marine ecosystems as primary producers in oceanic gyres, as agents of biological nitrogen fixation, and, in modified form, as the plastids of marine algae.[5]
Early photosynthetic systems, such as those from green and purple sulfur and green and purple nonsulfur bacteria, are thought to have been anoxygenic, using various molecules as electron donors. Green and purple sulfur bacteria are thought to have used hydrogen and sulfur as an electron donor. Green nonsulfur bacteria used various amino and other organic acids. Purple nonsulfur bacteria used a variety of nonspecific organic molecules.
Fossils of what are thought to be filamentous photosynthetic organisms have been dated at 3.4 billion years old.[6][7]
The main source of oxygen in the atmosphere is oxygenic photosynthesis, and its first appearance is sometimes referred to as the oxygen catastrophe. Geological evidence suggests that oxygenic photosynthesis, such as that in cyanobacteria, became important during the Paleoproterozoic era around 2 billion years ago. Modern photosynthesis in plants and most photosynthetic prokaryotes is oxygenic. Oxygenic photosynthesis uses water as an electron donor, which is oxidized to molecular oxygen (O
2) in the photosynthetic reaction center.
Symbiosis and the origin of chloroplasts
Several groups of animals have formed symbiotic relationships with photosynthetic algae. These are most common in corals, sponges and sea anemones. It is presumed that this is due to the particularly simple body plans and large surface areas of these animals compared to their volumes.[8] In addition, a few marine mollusks Elysia viridis and Elysia chlorotica also maintain a symbiotic relationship with chloroplasts they capture from the algae in their diet and then store in their bodies. This allows the mollusks to survive solely by photosynthesis for several months at a time.[9][10] Some of the genes from the plant cell nucleus have even been transferred to the slugs, so that the chloroplasts can be supplied with proteins that they need to survive.[11]
An even closer form of symbiosis may explain the origin of chloroplasts. Chloroplasts have many similarities with photosynthetic bacteria, including a circular chromosome, prokaryotic-type ribosomes, and similar proteins in the photosynthetic reaction center.[12][13] The endosymbiotic theory suggests that photosynthetic bacteria were acquired (by endocytosis) by early eukaryotic cells to form the first plant cells. Therefore, chloroplasts may be photosynthetic bacteria that adapted to life inside plant cells. Like mitochondria, chloroplasts still possess their own DNA, separate from the nuclear DNA of their plant host cells and the genes in this chloroplast DNA resemble those in cyanobacteria.[14] DNA in chloroplasts codes for redox proteins such as photosynthetic reaction centers. The CoRR Hypothesis proposes that this Co-location is required for Redox Regulation.
A 2010 study by researchers at Tel Aviv University discovered that the Oriental hornet (Vespa orientalis) converts sunlight into electric power using a pigment called xanthopterin. This is the first scientific evidence of a member of the animal kingdom engaging in photosynthesis.[15]
Evolution of photosynthetic pathways
Photosynthesis is not quite as simple as adding water to CO2 to produce sugars and oxygen. A complex chemical pathway is involved, facilitated along the way by a range of enzymes and co-enzymes. The enzyme RuBisCO is responsible for "fixing" CO2 – that is, it attaches it to a carbon-based molecule to form a sugar, which can be used by the plant, releasing an oxygen molecule along the way. However, the enzyme is notoriously inefficient, and just as effectively will also fix oxygen instead of CO2 in a process called photorespiration. This is energetically costly as the plant has to use energy to turn the products of photorespiration back into a form that can react with CO2.
Concentrating carbon
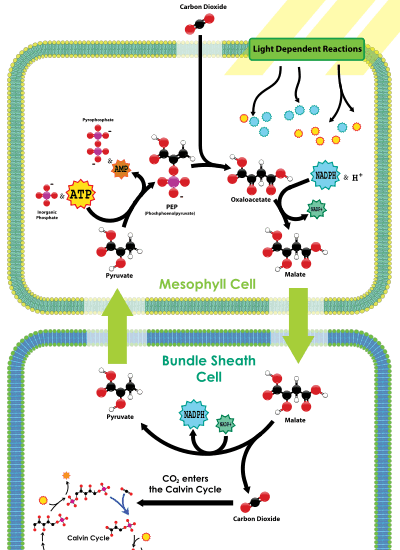
The C4 metabolic pathway is a valuable recent evolutionary innovation in plants, involving a complex set of adaptive changes to physiology and gene expression patterns.[16] About 7600 species of plants use C4 carbon fixation, which represents about 3% of all terrestrial species of plants. All these 7600 species are angiosperms.
C4 plants evolved carbon concentrating mechanisms. These work by increasing the concentration of CO2 around RuBisCO, thereby facilitating photosynthesis and decreasing photorespiration. The process of concentrating CO2 around RuBisCO requires more energy than allowing gases to diffuse, but under certain conditions – i.e. warm temperatures (>25 °C), low CO2 concentrations, or high oxygen concentrations – pays off in terms of the decreased loss of sugars through photorespiration.
One type of C4 metabolism employs a so-called Kranz anatomy. This transports CO2 through an outer mesophyll layer, via a range of organic molecules, to the central bundle sheath cells, where the CO2 is released. In this way, CO2 is concentrated near the site of RuBisCO operation. Because RuBisCO is operating in an environment with much more CO2 than it otherwise would be, it performs more efficiently.
A second mechanism, CAM photosynthesis, is a carbon fixation pathway that evolved in some plants as an adaptation to arid conditions.[17][18] The most important benefit of CAM to the plant is the ability to leave most leaf stomata closed during the day.[19] This reduces water loss due to evapotranspiration. The stomata open at night to collect CO2, which is stored as the four-carbon acid malate, and then used during photosynthesis during the day. The pre-collected CO2 is concentrated around the enzyme RuBisCO, increasing photosynthetic efficiency. More CO2 is then harvested from the atmosphere when stomata open, during the cool, moist nights, reducing water loss.
CAM has evolved convergently many times.[20] It occurs in 16,000 species (about 7% of plants), belonging to over 300 genera and around 40 families, but this is thought to be a considerable underestimate.[21] It is found in quillworts (relatives of club mosses), in ferns, and in gymnosperms, but the great majority of plants using CAM are angiosperms (flowering plants).
Evolutionary record
These two pathways, with the same effect on RuBisCO, evolved a number of times independently – indeed, C4 alone arose 62 times in 18 different plant families. A number of 'pre-adaptations' seem to have paved the way for C4, leading to its clustering in certain clades: it has most frequently been innovated in plants that already had features such as extensive vascular bundle sheath tissue.[22] Many potential evolutionary pathways resulting in the C4 phenotype are possible and have been characterised using Bayesian inference,[16] confirming that non-photosynthetic adaptations often provide evolutionary stepping stones for the further evolution of C4.

The C4 construction is most famously used by a subset of grasses, while CAM is employed by many succulents and cacti. The trait appears to have emerged during the Oligocene, around 25 to 32 million years ago;[23] however, they did not become ecologically significant until the Miocene, 6 to 7 million years ago.[24] Remarkably, some charcoalified fossils preserve tissue organised into the Kranz anatomy, with intact bundle sheath cells,[25] allowing the presence C4 metabolism to be identified without doubt at this time. Isotopic markers are used to deduce their distribution and significance.
C3 plants preferentially use the lighter of two isotopes of carbon in the atmosphere, 12C, which is more readily involved in the chemical pathways involved in its fixation. Because C4 metabolism involves a further chemical step, this effect is accentuated. Plant material can be analysed to deduce the ratio of the heavier 13C to 12C. This ratio is denoted δ13C. C3 plants are on average around 14‰ (parts per thousand) lighter than the atmospheric ratio, while C4 plants are about 28‰ lighter. The δ13C of CAM plants depends on the percentage of carbon fixed at night relative to what is fixed in the day, being closer to C3 plants if they fix most carbon in the day and closer to C4 plants if they fix all their carbon at night.[26]
It is troublesome procuring original fossil material in sufficient quantity to analyse the grass itself, but fortunately there is a good proxy: horses. Horses were globally widespread in the period of interest, and browsed almost exclusively on grasses. There's an old phrase in isotope palæontology, "you are what you eat (plus a little bit)" – this refers to the fact that organisms reflect the isotopic composition of whatever they eat, plus a small adjustment factor. There is a good record of horse teeth throughout the globe, and their δ13C has been measured. The record shows a sharp negative inflection around 6 to 7 million years ago, during the Messinian, and this is interpreted as the rise of C4 plants on a global scale.[24]
When is C4 an advantage?
While C4 enhances the efficiency of RuBisCO, the concentration of carbon is highly energy intensive. This means that C4 plants only have an advantage over C3 organisms in certain conditions: namely, high temperatures and low rainfall. C4 plants also need high levels of sunlight to thrive.[27] Models suggest that, without wildfires removing shade-casting trees and shrubs, there would be no space for C4 plants.[28] But, wildfires have occurred for 400 million years – why did C4 take so long to arise, and then appear independently so many times? The Carboniferous period (~300 million years ago) had notoriously high oxygen levels – almost enough to allow spontaneous combustion[29] – and very low CO2, but there is no C4 isotopic signature to be found. And there doesn't seem to be a sudden trigger for the Miocene rise.
During the Miocene, the atmosphere and climate were relatively stable. If anything, CO2 increased gradually from 14 to 9 million years ago before settling down to concentrations similar to the Holocene.[30] This suggests that it did not have a key role in invoking C4 evolution.[23] Grasses themselves (the group which would give rise to the most occurrences of C4) had probably been around for 60 million years or more, so had had plenty of time to evolve C4,[31][32] which, in any case, is present in a diverse range of groups and thus evolved independently. There is a strong signal of climate change in South Asia;[23] increasing aridity – hence increasing fire frequency and intensity – may have led to an increase in the importance of grasslands.[33] However, this is difficult to reconcile with the North American record.[23] It is possible that the signal is entirely biological, forced by the fire- (and elephant?)-[34] driven acceleration of grass evolution – which, both by increasing weathering and incorporating more carbon into sediments, reduced atmospheric CO2 levels.[34] Finally, there is evidence that the onset of C4 from 9 to 7 million years ago is a biased signal, which only holds true for North America, from where most samples originate; emerging evidence suggests that grasslands evolved to a dominant state at least 15Ma earlier in South America.
See also
References
- ↑ Olson JM (May 2006). "Photosynthesis in the Archean era". Photosyn. Res. 88 (2): 109–17. doi:10.1007/s11120-006-9040-5. PMID 16453059.
- ↑ Cardona T; Murray JW; Rutherford, AW (May 2015). "Origin and Evolution of Water Oxidation before the Last Common Ancestor of the Cyanobacteria". Molecular Biology and Evolution. 32 (5): 1310–1328. doi:10.1093/molbev/msv024. PMC 4408414
 . PMID 25657330.
. PMID 25657330. - ↑ Akiko Tomitani (April 2006). "The evolutionary diversification of cyanobacteria: Molecular–phylogenetic and paleontological perspectives". PNAS. 103 (14): 5442–5447. Bibcode:2006PNAS..103.5442T. doi:10.1073/pnas.0600999103. PMC 1459374. PMID 16569695.
- ↑ "Cyanobacteria: Fossil Record". Ucmp.berkeley.edu. Retrieved 2010-08-26.
- ↑ Herrero A, Flores E (2008). The Cyanobacteria: Molecular Biology, Genomics and Evolution (1st ed.). Caister Academic Press. ISBN 978-1-904455-15-8.
- ↑ Photosynthesis got a really early start, New Scientist, 2 October 2004
- ↑ Revealing the dawn of photosynthesis, New Scientist, 19 August 2006
- ↑ Venn AA, Loram JE, Douglas AE (2008). "Photosynthetic symbioses in animals". J. Exp. Bot. 59 (5): 1069–80. doi:10.1093/jxb/erm328. PMID 18267943.
- ↑ Rumpho ME, Summer EJ, Manhart JR (May 2000). "Solar-powered sea slugs. Mollusc/algal chloroplast symbiosis". Plant Physiol. 123 (1): 29–38. doi:10.1104/pp.123.1.29. PMC 1539252. PMID 10806222.
- ↑ Muscatine L, Greene RW (1973). "Chloroplasts and algae as symbionts in molluscs". Int. Rev. Cytol. International Review of Cytology. 36: 137–69. doi:10.1016/S0074-7696(08)60217-X. ISBN 9780123643360. PMID 4587388.
- ↑ Rumpho ME, Worful JM, Lee J, Kannan K, Tyler MS, Bhattacharya D, Moustafa A, Manhart JR (November 2008). "Horizontal gene transfer of the algal nuclear gene psbO to the photosynthetic sea slug Elysia chlorotica". Proc. Natl. Acad. Sci. U.S.A. 105 (46): 17867–71. Bibcode:2008PNAS..10517867R. doi:10.1073/pnas.0804968105. PMC 2584685. PMID 19004808.
- ↑ Douglas SE (December 1998). "Plastid evolution: origins, diversity, trends". Curr. Opin. Genet. Dev. 8 (6): 655–61. doi:10.1016/S0959-437X(98)80033-6. PMID 9914199.
- ↑ Reyes-Prieto A, Weber AP, Bhattacharya D (2007). "The origin and establishment of the plastid in algae and plants". Annu. Rev. Genet. 41: 147–68. doi:10.1146/annurev.genet.41.110306.130134. PMID 17600460.
- ↑ Raven JA, Allen JF (2003). "Genomics and chloroplast evolution: what did cyanobacteria do for plants?". Genome Biol. 4 (3): 209. doi:10.1186/gb-2003-4-3-209. PMC 153454. PMID 12620099.
- ↑ Plotkin, M.; Hod, I.; Zaban, A.; Boden, S. A.; Bagnall, D. M.; Galushko, D.; Bergman, D. J. (2010). "Solar energy harvesting in the epicuticle of the oriental hornet (Vespa orientalis)". Naturwissenschaften. 97 (12): 1067–1076. Bibcode:2010NW.....97.1067P. doi:10.1007/s00114-010-0728-1. PMID 21052618.
- 1 2 Williams BP, Johnston IG, Covshoff S, Hibberd JM (September 2013). "Phenotypic landscape inference reveals multiple evolutionary paths to C₄ photosynthesis". eLife. 2: e00961. doi:10.7554/eLife.00961. PMID 24082995.
- ↑ C.Michael Hogan. 2011. Respiration. Encyclopedia of Earth. Eds. Mark McGinley & C.J.cleveland. National council for Science and the Environment. Washington DC
- ↑ Herrera, A. (2008), "Crassulacean acid metabolism and fitness under water deficit stress: if not for carbon gain, what is facultative CAM good for?", Annals of Botany, 103 (4): 645–653, doi:10.1093/aob/mcn145
- ↑ Ting, I P (1985). "Crassulacean Acid Metabolism". Annual Review of Plant Physiology. 36 (1): 595–622. doi:10.1146/annurev.pp.36.060185.003115.
- ↑ Keeley, Jon E.; Rundel, Philip W. (2003). "Evolution of CAM and C4 Carbon‐Concentrating Mechanisms" (PDF). International Journal of Plant Sciences. 164 (S3): S55. doi:10.1086/374192.
- ↑ Dodd, A. N.; Borland, A. M.; Haslam, R. P.; Griffiths, H.; Maxwell, K. (2002). "Crassulacean acid metabolism: plastic, fantastic". Journal of Experimental Botany. 53 (369): 569–580. doi:10.1093/jexbot/53.369.569. PMID 11886877.
- ↑ Christin, P. -A.; Osborne, C. P.; Chatelet, D. S.; Columbus, J. T.; Besnard, G.; Hodkinson, T. R.; Garrison, L. M.; Vorontsova, M. S.; Edwards, E. J. (2012). "Anatomical enablers and the evolution of C4 photosynthesis in grasses". Proceedings of the National Academy of Sciences. 110 (4): 1381–1386. Bibcode:2013PNAS..110.1381C. doi:10.1073/pnas.1216777110.
- 1 2 3 4 Osborne, C.P.; Beerling, D.J. (2006). "Review. Nature's green revolution: the remarkable evolutionary rise of C4 plants" (PDF). Philosophical Transactions of the Royal Society B. 361 (1465): 173–194. doi:10.1098/rstb.2005.1737. PMC 1626541. PMID 16553316. Retrieved 2008-02-11.
- 1 2 Retallack, G. J. (1 August 1997). "Neogene Expansion of the North American Prairie". PALAIOS. 12 (4): 380–390. doi:10.2307/3515337. ISSN 0883-1351. JSTOR 3515337.
- ↑ Thomasson, J.R.; Nelson, M.E.; Zakrzewski, R.J. (1986). "A Fossil Grass (Gramineae: Chloridoideae) from the Miocene with Kranz Anatomy". Science. 233 (4766): 876–878. Bibcode:1986Sci...233..876T. doi:10.1126/science.233.4766.876. PMID 17752216.
- ↑ O'Leary, Marion (May 1988). "Carbon Isotopes in Photosynthesis". BioScience. 38 (5): 328–336. doi:10.2307/1310735. JSTOR 1310735.
- ↑ Osborne, P.; Freckleton, P. (Feb 2009). "Ecological selection pressures for C4 photosynthesis in the grasses". Proceedings. Biological sciences / the Royal Society. 276 (1663): 1753–1760. doi:10.1098/rspb.2008.1762. ISSN 0962-8452. PMC 2674487. PMID 19324795.
- ↑ Bond, W.J.; Woodward, F.I.; Midgley, G.F. (2005). "The global distribution of ecosystems in a world without fire". New Phytologist. 165 (2): 525–538. doi:10.1111/j.1469-8137.2004.01252.x. PMID 15720663.
- ↑ Above 35% atmospheric oxygen, the spread of fire is unstoppable. Many models have predicted higher values and had to be revised, because there was not a total extinction of plant life.
- ↑ Pagani, M.; Zachos, J.C.; Freeman, K.H.; Tipple, B.; Bohaty, S. (2005). "Marked Decline in Atmospheric Carbon Dioxide Concentrations During the Paleogene". Science. 309 (5734): 600–603. Bibcode:2005Sci...309..600P. doi:10.1126/science.1110063. PMID 15961630.
- ↑ Piperno, D.R.; Sues, H.D. (2005). "Dinosaurs Dined on Grass". Science. 310 (5751): 1126–8. doi:10.1126/science.1121020. PMID 16293745.
- ↑ Prasad, V.; Stroemberg, C.A.E.; Alimohammadian, H.; Sahni, A. (2005). "Dinosaur Coprolites and the Early Evolution of Grasses and Grazers". Science. 310 (5751): 1177–1180. Bibcode:2005Sci...310.1177P. doi:10.1126/science.1118806. PMID 16293759.
- ↑ Keeley, J.E.; Rundel, P.W. (2005). "Fire and the Miocene expansion of C4 grasslands". Ecology Letters. 8 (7): 683–690. doi:10.1111/j.1461-0248.2005.00767.x.
- 1 2 Retallack, G.J. (2001). "Cenozoic Expansion of Grasslands and Climatic Cooling". The Journal of Geology. 109 (4): 407–426. Bibcode:2001JG....109..407R. doi:10.1086/320791.