Polydnavirus
| Polydnavirus
| |
|---|---|
| Virus classification | |
| Group: | Group I (dsDNA) |
| Family: | Polydnaviridae |
| Genera | |

A polydnavirus /pɒˈlɪdnəvaɪərəs/ (PDV) is a member of the family Polydnaviridae of insect viruses. There are currently 53 species in this family, divided among 2 genera. Polydnaviruses form a symbiotic relationship with parasitoid wasps (ichnoviruses occur in ichneumonid wasps species and bracoviruses in braconid wasps), but these wasps are themselves parasites of lepidoptera.[1][2] Little or no sequence homology exists between BV and IV, suggesting that the two genera evolved independently.
Taxonomy
Group: dsDNA
- Family: Polydnaviridae
- Genus: Bracovirus
- Apanteles crassicornis bracovirus
- Apanteles fumiferanae bracovirus
- Ascogaster argentifrons bracovirus
- Ascogaster quadridentata bracovirus
- Cardiochiles nigriceps bracovirus
- Chelonus altitudinis bracovirus
- Chelonus blackburni bracovirus
- Chelonus inanitus bracovirus
- Chelonus insularis bracovirus
- Chelonus nr. curvimaculatus bracovirus
- Chelonus texanus bracovirus
- Cotesia congregata bracovirus
- Cotesia flavipes bracovirus
- Cotesia glomerata bracovirus
- Cotesia hyphantriae bracovirus
- Cotesia kariyai bracovirus
- Cotesia marginiventris bracovirus
- Cotesia melanoscela bracovirus
- Cotesia rubecula bracovirus
- Cotesia schaeferi bracovirus
- Diolcogaster facetosa bracovirus
- Glyptapanteles flavicoxis bracovirus
- Glyptapanteles indiensis bracovirus
- Glyptapanteles liparidis bracovirus
- Hypomicrogaster canadensis bracovirus
- Hypomicrogaster ectdytolophae bracovirus
- Microplitis croceipes bracovirus
- Microplitis demolitor bracovirus
- Phanerotoma flavitestacea bracovirus
- Pholetesor ornigis bracovirus
- Protapanteles paleacritae bracovirus
- Tranosema rostrale bracovirus
- Genus: Ichnovirus
- Campoletis aprilis ichnovirus
- Campoletis flavicincta ichnovirus
- Campoletis sonorensis ichnovirus
- Casinaria arjuna ichnovirus
- Casinaria forcipata ichnovirus
- Casinaria infesta ichnovirus
- Diadegma acronyctae ichnovirus
- Diadegma interruptum ichnovirus
- Diadegma terebrans ichnovirus
- Enytus montanus ichnovirus
- Eriborus terebrans ichnovirus
- Glypta fumiferanae ichnovirus
- Hyposoter annulipes ichnovirus
- Hyposoter exiguae ichnovirus
- Hyposoter fugitivus ichnovirus
- Hyposoter lymantriae ichnovirus
- Hyposoter pilosulus ichnovirus
- Hyposoter rivalis ichnovirus
- Olesicampe benefactor ichnovirus
- Olesicampe geniculatae ichnovirus
- Synetaeris tenuifemur ichnovirus
Structure
Viruses in Polydnaviridae are enveloped, with prolate ellipsoid and cylindrical geometries. Genomes are circular and segmented, composed of multiple segments of double-stranded, superhelical DNA packaged in capsid proteins. They are around 2.0–31kb in length.[1]
| Genus | Structure | Symmetry | Capsid | Genomic Arrangement | Genomic Segmentation |
|---|---|---|---|---|---|
| Ichnovirus | Prolate ellipsoid | Enveloped | Circular | Segmented | |
| Bracovirus | Prolate ellipsoid | Enveloped | Circular | Segmented |
Life Cycle
Viral replication is nuclear. DNA-templated transcription is the method of transcription. The virus exits the host cell by nuclear pore export. Parasitoid wasps serve as hosts, but these wasps are themselves parasites of lepidoptera. The wasp injects one or more eggs into its host along with a quantity of virus. The virus and wasp are in a symbiotic relationship: expression of viral genes prevents the wasp's host's immune system from killing the wasp's injected egg and causes other physiological alterations that ultimately cause the parasitized host to die. Transmission routes are parental.[1]
| Genus | Host Details | Tissue Tropism | Entry Details | Release Details | Replication Site | Assembly Site | Transmission |
|---|---|---|---|---|---|---|---|
| Ichnovirus | Parasitoid wasps (Ichneumonidae) | Hemocytes; fat bodies | Unknown | Lysis; budding | Nucleus | Nucleus | Unknown |
| Bracovirus | Parasitoid wasps (Braconidae) | Hemocytes; fat bodies | Unknown | Lysis; budding | Nucleus | Nucleus | Unknown |
Biology
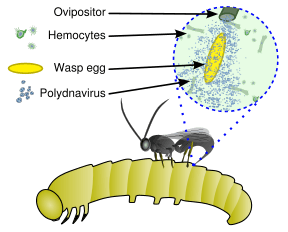
These viruses are part of a unique biological system consisting of an endoparasitic wasp (parasitoid), an insect (usually lepidopteran) larva, and the virus. The full genome of the virus is integrated into the genome of the wasp and the virus only replicates in a particular part of the ovary, called the calyx, of pupal and adult female wasps. The virus is injected along with the wasp egg into the body cavity of a lepidopteran host caterpillar and infects cells of the caterpillar. The infection does not lead to replication of new viruses, rather it affects the caterpillar's immune system. Without the virus infection, phagocytic hemocytes (blood cells) will encapsulate and kill the wasp egg but the immune suppression caused by the virus allows for survival of the wasp egg, leading to hatching and complete development of the immature wasp in the caterpillar. Additionally, genes expressed from the polydnavirus in the parasitised host alter host development and metabolism to be beneficial for the growth and survival of the parasitoid larva. Thus the virus and wasp have a symbiotic (mutualistic) relationship.[3][4]
Characteristics
Both genera of PDV share certain characteristics:
- the virus particles of each contain multiple segments of dsDNA (double-strand, or "normal" DNA, as contrasted with positive or negative-sense single-strand DNA or RNA, as found in some other viruses) with each segment containing only part of the full genome (much like chromosomes in eukaryotic organisms)
- the genome of the virus has eukaryotic characteristic such as the presence of introns and a low coding density
- the genome of each virus is integrated into the host wasp genome
- the genome is organized in several multiple members genes families (which are different between Bracoviruses and Ichnoviruses)
- the virus particles are only replicated (produced) in specific cell types in the female wasp's reproductive organs
However, the morphology of the two genera are different when observed by electron microscopy. Ichnoviruses tend to be ovoid (egg-shaped) while bracoviruses are short rods. The virions of Bracoviruses are released by cell lysis whereas virions of Ichnoviruses are released by budding.
Evolution
Nucleic acid analysis suggests a very long association of the viruses with the wasps (greater than 70 million years).
Two proposals have been advanced for how the wasp/virus association developed. The first suggests that the virus is derived from wasp genes. Many parasitoids that do not use PDVs inject proteins that provide many of the same functions, that is, a suppression of the immune response to the parasite egg. In this model, the braconid and ichneumonid wasps packaged genes for these functions into the viruses—essentially creating a gene-transfer system that results in the caterpillar producing the immune-suppressing factors. In this scenario, the PDV structural proteins (capsids) were probably "borrowed" from existing viruses.
The alternative proposal suggests that ancestral wasps developed a beneficial association with an existing virus that eventually led to the integration of the virus into the wasp’s genome. Following integration, the genes responsible for virus replication and the capsids were (eventually) no longer included in the PDV genome. This hypothesis is supported by the distinct morphology differences between IV and BV, suggesting different ancestral viruses for the two families. The IV have remarkable similarities to ascoviruses[5] while BV may have evolved from a nudivirus ~100 million years ago.[6]
The two groups of viruses in the family are not in fact phylogenetically related suggesting that this taxon may need revision.[7]
Effect on host immunity
In the host, several mechanisms of the insect immune system can be triggered when the wasp lays its eggs and when the parasitic wasp is developing.
When a large body (wasp egg or small particle used experimentally) is introduced into an insect's body, the classic immune reaction is the encapsulation by the hematocytes. An encapsulated body can also be melanised in order to asphyxiate it, thanks to another type of hemocyte, which uses the phenoloxidase pathway to produce melanin. Small particles can be phagocytosed, and macrophage cells can then be also melanised in a nodule. Finally, insects can also respond with production of antiviral peptides.[8]
PolyDNAvirus protect the hymenopteran larvae from the host immune system, acting at different levels.
- First they can disable or destroy hematocytes. The polyDNAvirus associated with Cotesia rubecula, code for a protein CrV1 that denatures actin filaments in hematocytes, so those cells become less able to move and adhere to the larvae.[8] Microplitis demolitor Bracovirus (MdBV) induce apoptosis of hematocytes, thanks to its gene PTP-H2.[8] It also decreases the adhesion capacity of hematocytes, thanks to its gene Glc1.8. The gene also inhibits phagocytosis.[9]
- PolyDNAvirus can also act on melanisation, MdBV interferes with the production of phenoloxidase.[10]
- Finally, polyDNAvirus can also produce viral ankyrins, that interfere with production of antiviral peptides.[11] In some Ichnoviruses, Vankyrin can also prevent apoptosis, the extreme reaction of a cell to block viral propagation.[12][13]
- The Ichnoviruses produce some proteins called vinnexins which have been recognized as homologous to the innexins of insects. They are responsible for the encoding of the structural units of the gap-junctions. These proteins may alter the intercellular communication which could explain the disruption of the encapsidation process.[14]
Virus like particles
Another strategy used by parasitoid Hymenoptera to protect their offspring is production of Virus Like Particles. VLPs are very close to viruses in their structure but they don't carry any nucleic acid. For example, Venturia canescens (Ichneumonidea) and Leptopilina sp. (Figitidaea) produce VLPs.
VLPs can be compared to PolyDNAvirus because they are secreted in the same way, and they both act to protect the larvae against the host's immune system. V. canescens-VLPs (VcVLP1, VcVLP2,VcNEP …) are produced in the calix cells before they go to the oviducts. But they are composed of proteins that are closer in structure and function to Hymenoptera's proteins than to viral proteins. This suggests that they don't have a viral origin, unlike polyDNAvirus.
VLPs protect the Hymenoptera larvae locally, whereas polyDNAvirus can have a more global effect. VLPs allow the larvae to escape the immune system: the larva is not recognised as harmful by its host, or the immune cells can't interact with it thanks to the VLPs.[15]
The wasp Leptopilina heterotoma secrete VLPs that are able to penetrate into the lamellocytes, thanks to specific receptors, and then modify their shape and surface properties so they become inefficient and the larvae are safe from encapsulation.[16]
Micro-RNA
MicroRNA are small RNA fragments produced in the host cells thanks to a specific enzymatic mechanism. They promote viral RNA destruction. MicroRNA attach to viral-RNA because they are complementary. Then the complex is recognised by an enzyme that destroys it. This phenomenon is known as PTGS (for post transcriptional gene silencing)[17]
It is interesting to consider the microRNA phenomenon in the polyDNAvirus context. Many hypotheses can be formulated :
- Braconidae carry nudivirus-related genes in their genome,[18] so they are may be able to produce microRNA against nudivirus, as an innate immunity.
- Wasps perhaps use microRNA to control the viral genes they carry.
- PolyDNAvirus can also use PTGS to interfere with the host's gene expression.
- PTGS is also used for organisms' development, using the same enzymes as antiviral gene silencing, so we can imagine that if the host uses PTGS against polyDNAvirus, perhaps it also affects its development.
See also
References
- 1 2 3 "Viral Zone". ExPASy. Retrieved 15 June 2015.
- 1 2 ICTV. "Virus Taxonomy: 2014 Release". Retrieved 15 June 2015.
- ↑ Webb, B. A. (1998). Polydnavirus biology, genome structure, and evolution. In Miller, L.K., Ball, L.A., Eds. The Insect Viruses. Plenum Publishing Corporation. pp. 105–139.
- ↑ Roossinck, M. J. (2011). Changes in population dynamics in mutualistic versus pathogenic viruses. Viruses 3: 12–19.
- ↑ Federici BA, Bideshi DK, Tan Y, Spears T, Bigot Y (2009) Ascoviruses: superb manipulators of apoptosis for viral replication and transmission. Curr Top Microbiol Immunol 328:171–196
- ↑ Herniou EA, Huguet E, Thézé J, Bézier A, Periquet G, Drezen JM (2013). "When parasitic wasps hijacked viruses: genomic and functional evolution of polydnaviruses". Philos Trans R Soc Lond B Biol Sci. 368 (1626): 20130051. doi:10.1098/rstb.2013.0051.
- ↑ Dupuy C, Huguet E, Drezen JM (2006). "Unfolding the evolutionary story of polydnaviruses". Virus Res. 117 (1): 81–89.
- 1 2 3 JM. Drezen; S. Savary; M. Poirier; G. Periquet (1999). "Polydnaviruses, viral entities domesticated by the parasitoid wasps". Virologie. 3 (1): 11–21.
- ↑ Markus Beck & Michael R. Strand (February 2005). "Glc1.8 from Microplitis demolitor Bracovirus Induces a Loss of Adhesion and Phagocytosis in Insect High Five and S2 Cells". Journal of Virology. 79 (3): 1861–1870. doi:10.1128/jvi.79.3.1861-1870.2005.
- ↑ Lu Zhiqiang; Beck Markus H.; Strand Michael R (Jul 2010). "The Viral Protein Egf1.0 Is a Dual Activity Inhibitor of Prophenoloxidase-activating Proteinases 1 and 3 from Manduca sexta". INSECT BIOCHEMISTRY AND MOLECULAR BIOLOGY. 40 (: 7).
- ↑ Bae Sungwoo; Kim Yonggyun (2009-09-01). "IkB genes encoded in Cotesia plutellae bracovirus suppress an antiviral response and enhance baculovirus pathogenicity against the diamondback moth, Plutella xylostella.". Journal of Invertebrate Pathology. 102 (1): 79–87. doi:10.1016/j.jip.2009.06.007. ISSN 0022-2011.
- ↑ Fath-Goodin A.; Kroemer J. A.; Webb B. A (Aug 2009). "The Campoletis sonorensis ichnovirus vankyrin protein P-vank-1 inhibits apoptosis in insect Sf9 cells". INSECT MOLECULAR BIOLOGY. 18 (4): 497–506. doi:10.1111/j.1365-2583.2009.00892.x.
- ↑ Clavijo G, Dorémus T, Ravallec M, Mannucci MA, Jouan V, Volkoff AN, Darboux I (2011). "Multigenic families in Ichnovirus: A tissue and host specificity study through expression analysis of vankyrins from Hyposoter didymator Ichnovirus". PLoS ONE. 6 (11): e27522. doi:10.1371/journal.pone.0027522.
- ↑ Marziano N.K.; Hasegawa D.K.; Phelan P.; Turnbull M.W. (Oct 2011). "Functional Interactions between Polydnavirus and Host Cellular Innexins". Journal of Virology. 85 (: 19): Pages: 10222–10229. doi:10.1128/jvi.00691-11.
- ↑ Annette Reineke; Sassan Asgari & Otto Schmidt (2006). "Evolutionary Origin of V enturia canescens Virus-Like Particles". Archives of Insect Biochemistry and Physiology. 61: 123–133. doi:10.1002/arch.20113.
- ↑ R. M. Rizki et T. M. Rizki, (November 1990). "Parasitoid virus-like particles destroy Drosophila cellular immunity". Proceedings of the National Academy of Sciences of the United States of America. 87: 8388–8392. doi:10.1073/pnas.87.21.8388.
- ↑ Li, H.W. & Ding, S.W. (2005). "Antiviral silencing in animals". FEBS Lett. 579: 5965–5973. doi:10.1016/j.febslet.2005.08.034. PMC 1350842
 . PMID 16154568.
. PMID 16154568. - ↑ Bézier A, Annaheim M, Herbinière J, Wetterwald C, Gyapay G, Bernard-Samain S, Wincker P, Roditi I, Heller M, Belghazi M, Pfister-Wilhem R, Periquet G, Dupuy C, Huguet E, Volkoff AN, Lanzrein B, Drezen JM (13 February 2009). "Polydnaviruses of braconid wasps derive from an ancestral nudivirus". Science. 13. 323 (5916): 926–30. doi:10.1126/science.1166788. PMID 19213916.
- ICTVdB Management (2006). 00.055. Polydnaviridae. In: ICTVdB—The Universal Virus Database, version 4. Büchen-Osmond, C. (Ed), Columbia University, New York, USA
- Fleming, J.-A. G. W. & Summers, M. D. (1991). "Polydnavirus DNA is integrated in the DNA of its parasitoid wasp host". Proceedings of the National Academy of Sciences. 88 (21): 9770–9774. doi:10.1073/pnas.88.21.9770. PMC 52802. PMID 1946402.
- Whitfield, J. B. (2002). "Estimating the age of the polydnavirus-braconid wasp symbiosis". Proceedings of the National Academy of Sciences. 99 (11): 7508–7513. doi:10.1073/pnas.112067199. PMC 124262. PMID 12032313.
- Espagne, E.; et al. (2004). "Genome Sequence of a Polydnavirus: Insights into Symbiotic Virus Evolution". Science. 306 (5694): 286–289. doi:10.1126/science.1103066. PMID 15472078.
- Webb, B. A.; et al. (2006). "Polydnavirus genomes reflect their dual roles as mutualists and pathogens". Virology. 347 (1): 160–174. doi:10.1016/j.virol.2005.11.010. PMID 16380146.
- Bernard-Samain, S., Wincker, P., IRoditi, I., Heller, M., Belghazi, M.,Pfister-Wilhem, R., Periquet, G., Dupuy,C., Huguet, E., Volkoff, A.N., Lanzrein, B., Drezen, J.M., Polydnaviruses of Braconid Wasps
External links
- http://research.biology.arizona.edu/mosquito/willott/507/polydnaviruses.html
- Polydnavirus DNA is Integrated in the DNA of Its Parasitoid Wasp Host—article from the Proceedings of the National Academy of Sciences.
- Abstract of the PNAS article.
- Genome Sequence of a Polydnavirus: Insights into Symbiotic Virus Evolution article from Science.
- Viralzone: Polydnaviridae
- ICTV