Visual phototransduction
Visual phototransduction is the sensory transduction of the visual system. It is a process by which light is converted into electrical signals in the rod cells, cone cells and photosensitive ganglion cells of the retina of the eye.This cycle was elucidated by George Wald(1906-1997) for which he received the Nobel Prize in 1967.It is so called "Wald's Visual Cycle" after him.
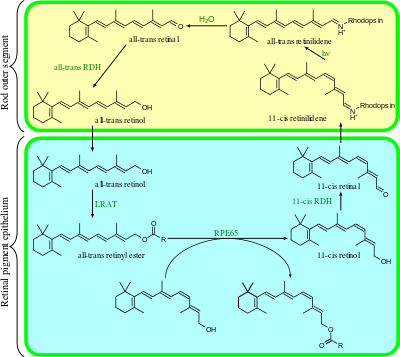
The visual cycle is the biological conversion of a photon into an electrical signal in the retina. This process occurs via G-protein coupled receptors called opsins which contain the chromophore 11-cis retinal. 11-cis retinal is covalently linked to the opsin receptor via Schiff base forming retinylidene protein. When struck by a photon, 11-cis retinal undergoes photoisomerization to all-trans retinal which changes the conformation of the opsin GPCR leading to signal transduction cascades which causes closure of cyclic GMP-gated cation channel, and hyperpolarization of the photoreceptor cell.
Following isomerization and release from the opsin protein, all-trans retinal is reduced to all-trans retinol and travels back to the retinal pigment epithelium to be "recharged". It is first esterified by lecithin retinol acyltransferase (LRAT) and then converted to 11-cis retinol by the isomerohydrolase RPE65. The isomerase activity of RPE65 has been shown; it is still uncertain whether it also acts as hydrolase. Finally, it is oxidized to 11-cis retinal before traveling back to the rod outer segment where it is again conjugated to an opsin to form new, functional visual pigment (rhodopsin).
Photoreceptors
The photoreceptor cells involved in vision are the rods and cones. These cells contain a chromophore (11-cis retinal, the aldehyde of Vitamin A1 and light-absorbing portion) bound to cell membrane protein, opsin. Rods deal with low light level and do not mediate color vision. Cones, on the other hand, can code the color of an image through comparison of the outputs of the three different types of cones. Each cone type responds best to certain wavelengths, or colors, of light because each type has a slightly different opsin. The three types of cones are L-cones, M-cones and S-cones that respond optimally to long wavelengths (reddish color), medium wavelengths (greenish color), and short wavelengths (bluish color) respectively. Humans have a trichromatic visual system consisting of three unique systems, rods, mid and long-wavelength sensitive (red and green) cones and short wavelength sensitive (blue) cones.[1]
Process

To understand the photoreceptor's behaviour to light intensities, it is necessary to understand the roles of different currents.
There is an ongoing outward potassium current through nongated K+-selective channels. This outward current tends to hyperpolarize the photoreceptor at around -70 mV (the equilibrium potential for K+).
There is also an inward sodium current carried by cGMP-gated sodium channels. This so-called 'dark current' depolarizes the cell to around -40 mV. Note that this is significantly more depolarized than most other neurons.
A high density of Na+-K+ pumps enables the photoreceptor to maintain a steady intracellular concentration of Na+ and K+.
In the dark
Photoreceptor cells are unusual cells in that they depolarize in response to absence of stimuli or scotopic conditions (darkness). In photopic conditions (light), photoreceptors hyperpolarize to a potential of -60mV. It is this 'switching off' that activates the next cell and sends an excitatory signal down the neural pathway.
In the dark, cGMP levels are high and keep cGMP-gated sodium channels open allowing a steady inward current, called the dark current. This dark current keeps the cell depolarized at about -40 mV.
The depolarization of the cell membrane in scotopic conditions opens voltage-gated calcium channels. An increased intracellular concentration of Ca2+ causes vesicles containing neurotransmitters, to merge with the cell membrane, therefore releasing the neurotransmitter into the synaptic cleft, an area between the end of one cell and the beginning of another neuron. The neurotransmitter released is glutamate, a neurotransmitter whose receptors are often excitatory.
In the cone pathway glutamate:
- Hyperpolarizes on-center bipolar cells. Glutamate that is released from the photoreceptors in the dark binds to metabotropic glutamate receptors (mGluR6), which, through a G-protein coupling mechanism, causes non-specific cation channels in the cells to close, thus hyperpolarizing the bipolar cell.
- Depolarizes off-center bipolar cells. Binding of glutamate to ionotropic glutamate receptors results in an inward cation current that depolarizes the bipolar cell.
In the light
- A light photon interacts with the retinal in a photoreceptor cell. The retinal undergoes isomerisation, changing from the 11-cis to all-trans configuration
- Retinal no longer fits into the opsin binding site.
- Opsin therefore undergoes a conformational change to metarhodopsin II.
- Metarhodopsin II is unstable and splits, yielding opsin and all-trans retinal.
- The opsin activates the regulatory protein transducin. This causes transducin to dissociate from its bound GDP, and bind GTP, then the alpha subunit of transducin dissociates from the beta and gamma subunits, with the GTP still bound to the alpha subunit.
- The alpha subunit-GTP complex activates phosphodiesterase or PDE.
- PDE breaks down cGMP to 5'-GMP. This lowers the concentration of cGMP and therefore the sodium channels close.
- Closure of the sodium channels causes hyperpolarization of the cell due to the ongoing efflux of potassium ions.
- Hyperpolarization of the cell causes voltage-gated calcium channels to close.
- As the calcium level in the photoreceptor cell drops, the amount of the neurotransmitter glutamate that is released by the cell also drops. This is because calcium is required for the glutamate-containing vesicles to fuse with cell membrane and release their contents.
- A decrease in the amount of glutamate released by the photoreceptors causes depolarization of On center bipolar cells (rod and cone On bipolar cells) and hyperpolarization of cone off-center bipolar cells.
Deactivation of the phototransduction cascade
GTPase Accelerating Protein (GAP) interacts with the alpha subunit of transducin, and causes it to hydrolyse its bound GTP to GDP, and thus halts the action of phosphodiesterase, stopping the transformation of cGMP to GMP.
Guanylate Cyclase Activating Protein (GCAP) is a calcium binding protein, and as the calcium levels in the cell have decreased, GCAP dissociates from its bound calcium ions, and interacts with Guanylate Cyclase, activating it. Guanylate Cyclase then proceeds to transform GTP to cGMP, replenishing the cell's cGMP levels and thus reopening the sodium channels that were closed during phototransduction.
Finally, Metarhodopsin II is deactivated. Recoverin, another calcium binding protein, is normally bound to Rhodopsin Kinase when calcium is present. When the calcium levels fall during phototransduction, the calcium dissociates from recoverin, and rhodopsin kinase is released, when it proceeds to phosphorylate metarhodopsin II, which decreases its affinity for transducin. Finally, arrestin, another protein, binds the phosphorylated metarhodopsin II, completely deactivating it. Thus, finally, phototransduction is deactivated, and the dark current and glutamate release is restored. It is this pathway, where Metarhodopsin II is phosphorylated and bound to arrestin and thus deactivated, which is thought to be responsible for the S2 component of dark adaptation. The S2 component represents a linear section of the dark adaptation function present at the beginning of dark adaptation for all bleaching intensities.
All-trans retinal is transported to the pigment epithelial cells to be reduced to all-trans retinol, the precursor to 11-cis retinal. This is then transported back to the rods. All-trans retinal cannot be synthesised by humans and must be supplied by vitamin A in the diet. Deficiency of all-trans retinal can lead to night blindness. This is part of the bleach and recycle process of retinoids in the photoreceptors and retinal pigment epithelium.
Phototransduction in invertebrates
Phototransduction process in invertebrates like the fruit fly is different from the vertebrates. Here, light induces the conformational change into Rhodopsin and converts it into meta-rhodopsin. This helps in dissociation of G -protein complex. Alpha sub-unit of this complex activates the PLC enzyme (PLC-beta) which hydrolyze the PIP2 into DAG. This hydrolysis leads to opening of TRP channels and influx of calcium.
References
- ↑ Ebrey, Thomas; Koutalos, Yiannis (January 2001). "Vertebrate Photoreceptors". Progress in Retinal and Eye Research. 20 (1): 49–94. doi:10.1016/S1350-9462(00)00014-8. Retrieved September 15, 2013.
- ↑ Leskov, Ilya; Klenchin,Handy,Whitlock,Govardovskii,Bownds,Lamb,Pugh,Arshavsky (September 2000). "The Gain of Rod Phototransduction: Reconciliation of Biochemical and Electrophysiological Measurements". Neuron. 27 (3): 525–537. doi:10.1016/S0896-6273(00)00063-5. Retrieved September 15, 2013.
- Moiseyev G, Chen Y, Takahashi Y, Wu BX, Ma JX. RPE65 is the isomerohydrolase in the retinoid visual cycle. Proc. Natl. Acad. Sci. 2005 Article.
- Jin M, Li S, Moghrabi WN, Sun H, Travis GH. Rpe65 is the retinoid isomerase in bovine retinal pigment epithelium. Cell. 2005 Article.
External links
- Visual pigments and visual transduction at med.utah.edu
- A General Overview on Visual Perception at brynmawr.edu
- Phototransduction at the US National Library of Medicine Medical Subject Headings (MeSH)