Zinc finger nuclease
Zinc-finger nucleases (ZFNs) are artificial restriction enzymes generated by fusing a zinc finger DNA-binding domain to a DNA-cleavage domain. Zinc finger domains can be engineered to target specific desired DNA sequences and this enables zinc-finger nucleases to target unique sequences within complex genomes. By taking advantage of endogenous DNA repair machinery, these reagents can be used to precisely alter the genomes of higher organisms. Alongside CRISPR/Cas9 and TALEN, ZFN is a prominent tool in the field of genome editing.
Domains
DNA-binding domain
The DNA-binding domains of individual ZFNs typically contain between three and six individual zinc finger repeats and can each recognize between 9 and 18 basepairs. If the zinc finger domains perfectly recognize a 3 basepair DNA sequence to generate a 3-finger array that can recognize a 9 basepair target site. Other procedures can utilize either 1-finger or 2-finger modules to generate zinc-finger arrays with six or more individual zinc fingers. The main drawback with this procedure is the specificities of individual zinc fingers can overlap and can depend on the context of the surrounding zinc fingers and DNA. Without methods to account for this "context dependence", the standard modular assembly procedure often fails unless it is used to recognize sequences of the form (GNN)N.[1]
Numerous selection methods have been used to generate zinc-finger arrays capable of targeting desired sequences. Initial selection efforts utilized phage display to select proteins that bound a given DNA target from a large pool of partially randomized zinc-finger arrays. More recent efforts have utilized yeast one-hybrid systems, bacterial one-hybrid and two-hybrid systems, and mammalian cells. A promising new method to select novel zinc-finger arrays utilizes a bacterial two-hybrid system and has been dubbed "OPEN" by its creators.[2] This system combines pre-selected pools of individual zinc fingers that were each selected to bind a given triplet and then utilizes a second round of selection to obtain 3-finger arrays capable of binding a desired 9-bp sequence. This system was developed by the Zinc-Finger Consortium as an alternative to commercial sources of engineered zinc-finger arrays.
(see: Zinc finger chimera for more info on zinc finger selection techniques)
DNA-cleavage domain
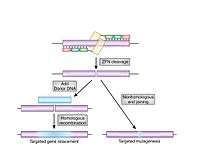
The non-specific cleavage domain from the type IIs restriction endonuclease FokI is typically used as the cleavage domain in ZFNs.[4] This cleavage domain must dimerize in order to cleave DNA[5] and thus a pair of ZFNs are required to target non-palindromic DNA sites. Standard ZFNs fuse the cleavage domain to the C-terminus of each zinc finger domain. In order to allow the two cleavage domains to dimerize and cleave DNA, the two individual ZFNs must bind opposite strands of DNA with their C-termini a certain distance apart. The most commonly used linker sequences between the zinc finger domain and the cleavage domain requires the 5' edge of each binding site to be separated by 5 to 7 bp.[6]
Several different protein engineering techniques have been employed to improve both the activity and specificity of the nuclease domain used in ZFNs. Directed evolution has been employed to generate a FokI variant with enhanced cleavage activity that the authors dubbed "Sharkey".[7] Structure-based design has also been employed to improve the cleavage specificity of FokI by modifying the dimerization interface so that only the intended heterodimeric species are active.[8][9][10][11]
Applications
Zinc finger nucleases are useful to manipulate the genomes of many plants and animals including arabidopsis,[12][13] tobacco,[14][15] soybean,[16] corn,[17] Drosophila melanogaster,[18] C. elegans,[19] Platynereis dumerilii,[20] sea urchin,[21] silkworm,[22] zebrafish,[23] frogs,[24] mice,[25] rats,[26] rabbits,[27] pigs,[28] cattle,[29] and various types of mammalian cells.[30] Zinc finger nucleases have also been used in a mouse model of haemophilia[31] and a clinical trial found CD4+ human T-cells with the CCR5 gene disrupted by zinc finger nucleases to be save as a potential treatment for HIV/AIDS.[32] ZFNs are also used to create a new generation of genetic disease models called isogenic human disease models.
Disabling an allele
ZFNs can be used to disable dominant mutations in heterozygous individuals by producing double-strand breaks (DSBs) in the DNA (see Genetic recombination) in the mutant allele, which will, in the absence of a homologous template, be repaired by non-homologous end-joining (NHEJ). NHEJ repairs DSBs by joining the two ends together and usually produces no mutations, provided that the cut is clean and uncomplicated. In some instances, however, the repair will be imperfect, resulting in deletion or insertion of base-pairs, producing frame-shift and preventing the production of the harmful protein.[33] Multiple pairs of ZFNs can also be used to completely remove entire large segments of genomic sequence.[34] To monitor the editing activity, a PCR of the target area will amplify both alleles and, if one contains an insertion, deletion, or mutation, it will result in a heteroduplex single-strand bubble that cleavage assays can easily detect. ZFNs have also been used to modify disease-causing alleles in triplet repeat disorders. Expanded CAG/CTG repeat tracts are the genetic basis for more than a dozen inherited neurological disorders including Huntington’s disease, myotonic dystrophy, and several spinocerebellar ataxias. It has been demonstrated in human cells that ZFNs can direct double-strand breaks (DSBs) to CAG repeats and shrink the repeat from long pathological lengths to short, less toxic lengths.[35]
Recently, a group of researchers have successfully applied the ZFN technology to genetically modify the gol pigment gene and the ntl gene in zebrafish embryo. Specific zinc-finger motifs were engineered to recognize distinct DNA sequences. The ZFN-encoding mRNA was injected into one-cell embryos and a high percentage of animals carried the desired mutations and phenotypes. Their research work demonstrated that ZFNs can specifically and efficiently create heritable mutant alleles at loci of interest in the germ line, and ZFN-induced alleles can be propagated in subsequent generations.
Similar research of using ZFNs to create specific mutations in zebrafish embryo has also been carried out by other research groups. The kdr gene in zebra fish encodes for the vascular endothelial growth factor-2 receptor. Mutagenic lesions at this target site was induced using ZFN technique by a group of researchers in US. They suggested that the ZFN technique allows straightforward generation of a targeted allelic series of mutants; it does not rely on the existence of species-specific embryonic stem cell lines and is applicable to other vertebrates, especially those whose embryos are easily available; finally, it is also feasible to achieve targeted knock-ins in zebrafish, therefore it is possible to create human disease models that are heretofore inaccessible.
Allele editing
ZFNs are also used to rewrite the sequence of an allele by invoking the homologous recombination (HR) machinery to repair the DSB using the supplied DNA fragment as a template. The HR machinery searches for homology between the damaged chromosome and the extra-chromosomal fragment and copies the sequence of the fragment between the two broken ends of the chromosome, regardless of whether the fragment contains the original sequence. If the subject is homozygous for the target allele, the efficiency of the technique is reduced since the undamaged copy of the allele may be used as a template for repair instead of the supplied fragment.
Gene therapy
The success of gene therapy depends on the efficient insertion of therapeutic genes at the appropriate chromosomal target sites within the human genome, without causing cell injury, oncogenic mutations or an immune response. The construction of plasmid vectors is simple and straightforward. Custom-designed ZFNs that combine the non-specific cleavage domain (N) of FokI endonuclease with zinc-finger proteins (ZFPs) offer a general way to deliver a site-specific DSB to the genome, and stimulate local homologous recombination by several orders of magnitude. This makes targeted gene correction or genome editing a viable option in human cells. Since ZFN-encoding plasmids could be used to transiently express ZFNs to target a DSB to a specific gene locus in human cells, they offer an excellent way for targeted delivery of the therapeutic genes to a pre-selected chromosomal site. The ZFN-encoding plasmid-based approach has the potential to circumvent all the problems associated with the viral delivery of therapeutic genes.[36] The first therapeutic applications of ZFNs are likely to involve ex vivo therapy using a patients own stem cells. After editing the stem cell genome, the cells could be expanded in culture and reinserted into the patient to produce differentiated cells with corrected functions. The initial targets will likely include the causes of monogenic diseases such as the IL2Rγ gene and the b-globin gene for gene correction and CCR5 gene for mutagenesis and disablement.[33]
Potential problems
Off-target cleavage
If the zinc finger domains are not specific enough for their target site or they do not target a unique site within the genome of interest, off-target cleavage may occur. Such off-target cleavage may lead to the production of enough double-strand breaks to overwhelm the repair machinery and, as a consequence, yield chromosomal rearrangements and/or cell death. Off-target cleavage events may also promote random integration of donor DNA.[33] Two separate methods have been demonstrated to decrease off-target cleavage for 3-finger ZFNs that target two adjacent 9-basepair sites.[37] Other groups use ZFNs with 4, 5 or 6 zinc fingers that target longer and presumably rarer sites and such ZFNs could theoretically yield less off-target activity. A comparison of a pair of 3-finger ZFNs and a pair of 4-finger ZFNs detected off-target cleavage in human cells at 31 loci for the 3-finger ZFNs and at 9 loci for the 4-finger ZFNs.[38] Whole genome sequencing of C. elegans modified with a pair of 5-finger ZFNs found only the intended modification and a deletion at a site "unrelated to the ZFN site" indicating this pair of ZFNs was capable of targeting a unique site in the C. elegans genome.[19]
Immunogenicity
As with many foreign proteins inserted into the human body, there is a risk of an immunological response against the therapeutic agent and the cells in which it is active. Since the protein will need to be expressed only transiently, however, the time over which a response may develop is short.[33] Liu et al. respectively target ZFNickases to the endogenous b-casein(CSN2) locus stimulates lysostaphin and human lysozyme gene addition by homology-directed repair and derive secrete lysostaphin cows.[39][40]
Prospects
The ability to precisely manipulate the genomes of plants, animals and insects has numerous applications in basic research, agriculture, and human therapeutics. Using ZFNs to modify endogenous genes has traditionally been a difficult task due mainly to the challenge of generating zinc finger domains that target the desired sequence with sufficient specificity. Improved methods of engineering zinc finger domains and the availability of ZFNs from a commercial supplier now put this technology in the hands of increasing numbers of researchers. Several groups are also developing other types of engineered nucleases including engineered homing endonucleases[41] [42] and nucleases based on engineered TAL effectors.[43][44] TAL effector nucleases (TALENs) are particularly interesting because TAL effectors appear to be very simple to engineer[45] [46] and TALENs can be used to target endogenous loci in human cells.[47] But to date no one has reported the isolation of clonal cell lines or transgenic organisms using such reagents. One type of ZFN, known as SB-728-T, has been tested for potential application in the treatment of HIV.[48]
Zinc-finger nickases
Zinc-finger nickases (ZFNickases) are created by inactivating the catalytic activity of one ZFN monomer in the ZFN dimer required for double-strand cleavage.[49] ZFNickases demonstrate strand-specific nicking activity in vitro and thus provide for highly specific single-strand breaks in DNA.[49] These SSBs undergo the same cellular mechanisms for DNA that ZFNs exploit, but they show a significantly reduced frequency of mutagenic NHEJ repairs at their target nicking site. This reduction provides a bias for HR-mediated gene modifications. ZFNickases can induce targeted HR in cultured human and livestock cells, although at lower levels than corresponding ZFNs from which they were derived because nicks can be repaired without genetic alteration.[39][50] A major limitation of ZFN-mediated gene modifications is the competition between NHEJ and HR repair pathways. Regardless of the presence of a DNA donor construct, both repair mechanisms can be activated following DSBs induced by ZFNs. Thus, ZFNickases is the first plausible attempt at engineering a method to favor the HR method of DNA repair as opposed to the error-prone NHEJ repair. By reducing NHEJ repairs, ZFNickases can thereby reduce the spectrum of unwanted off-target alterations. The ease by which ZFNickases can be derive from ZFNs provides a great platform for further studies regarding the optimization of ZFNickases and possibly increasing their levels of targeted HR while still maintain their reduced NHEJ frequency.
Zinc finger nuclease treatment of HIV
Since antiretroviral therapy requires a lifelong treatment regimen, research to find more permanent cures for HIV infection is currently underway.[51] It is possible to synthesize zinc finger nucleotides with zinc finger components that selectively (almost selectively) bind to specific portions of DNA. Conceptually, targeting and editing could focus on host cellular co-receptors for HIV or on proviral HIV DNA.
Host cellular co-receptors for HIV
It has also been observed that 20% of the Caucasian population possess a mutation, called CCR5-Δ32 (frequency of 0.0808 for homozygous allele), that prevents the CCR5 chemokine receptor protein, which is the main means of viral access into the cell, from being expressed on the surface of their CD4+ T-cells.[52][53][54][55][56] Individuals who are homozygous for this mutation are immune to HIV strains that utilize the CCR5 receptor in order to gain access to the cell while those who are heterozygous for this mutation have been found to have reduced plasma viral load in addition to a delayed progression to AIDS.[57][58] By combining these facts, researchers have proposed a novel method of treatment for HIV. This method attempts to treat the infection by disrupting the CCR5 gene, such as introducing the CCR5-Δ32 mutation using a recombinant adenoviral vector or forcing DNA repair by nonhomologous end joining, which is prone to error and results in a non-functional gene. As a consequence, resulting in the expression of nonfunctional CCR5 co-receptors on CD4+ T cells, providing immunity against infection.[3][57][59][60]
The zinc finger nucleases that have been synthesized for this treatment are manufactured by combining FokI Type II restriction endonucleases with engineered zinc fingers.[3][61] The number of zinc fingers attached to the endonuclease controls the specificity of the ZFN since they are engineered to preferentially bind to specific base sequences in DNA. Each ZFN is made up of multiple zinc fingers and one nuclease enzyme.[3]
Proviral HIV DNA
A recent and unique application of ZFN-technology to treat HIV has emerged whose focus is to target not the host genome, but rather proviral HIV DNA, for mutagenesis.[62] The authors of this work have drawn their inspiration from the innate defense mechanism against bacteria-infecting-viruses called bacteriophages, present amongst those bacteria endowed with restriction modification (R-M) systems. These bacteria secrete a restriction enzyme (REase) that recognizes and repetitively cleaves around palindromic sequences within the xenogenic DNAs of the bacteriophages or simply phages, until the same is disabled. Further support for this approach resides in the fact that, the human genome comprises in large part remnants of retroviral genomes that have been inactivated by several mechanisms, some of whose action resembles that of ZFN. It should not be surprising, therefore, that the initial work leading to the application of ZFN technology in this manner revolved around and involved the isolation and testing of HIV/SIV targeting bacteria-derived REases, whose non-specificity (due to their short recognition sequences) unfortunately, rendered them toxic to the host genome. The latter-potential host-genome toxicity posed by the raw bacteria-derived REases limited their application to ex-vivo modalities for HIV prevention, namely synthetic or live microbicides. Subsequently, however, the unique specificity offered by ZFNs was quickly recognized and harnessed, paving way for a novel strategy for attacking HIV in-vivo (through target mutagenesis of proviral HIV DNA) that is similar to the manner by which bacteria equipped with R-M systems do, to disable the foreign DNAs of in-coming phage-genomes. Because latent proviral HIV DNA resident in resting memory CD4 cells forms the major barrier to the eradication of HIV by highly active antiviral therapy (HAART), it is speculated that this approach may offer a 'functional cure" for HIV. Both ex-vivo (manipulation of stem or autologous T cell precursors) and in-vivo delivery platforms are being explored. It is also hoped that, when applied to non-HIV infected persons, this strategy could offer a genomic vaccine against HIV and other viruses. Similar work is ongoing for high-risk HPVs (with the intent of reversing cervical neoplasia) [63] as well as with HSV-2 (with the goal of achieving a complete cure for genital herpes) [64][65][66][67][68][69][70][71][72][73][74]
Zinc finger binding
The FokI catalytic domain must dimerize in order to cleave the DNA at the targeted site, and requires there to be two adjacent zinc finger nucleases (see picture), which independently bind to a specific codon at the correct orientation and spacing. As a result, the two binding events from the two zinc finger nuclease enables specific DNA targeting.[75] Specificity of genome editing is important in order for the zinc finger nuclease to be a successful application. The consequence of off-targeting cleavage can lead to a decrease in efficiency of the on-target modification in addition to other unwanted changes.[75]
The exact constitution of the ZFNs that are to be used to treat HIV is still unknown. The binding of ZFNs for the alteration of the Zif268 genelink, however, has been well-studied and is outlined below in order to illustrate the mechanism by which the zinc finger domain of ZFNs bind to DNA.[76][77]
The amino terminus of the alpha helix portion of zinc fingers targets the major grooves of the DNA helix and binds near the CCR5 gene positioning FokI in a suitable location for DNA cleavage.[3][76][77]
Zinc fingers are repeated structural protein motifs with DNA recognition function that fit in the major grooves of DNA.[76] Three zinc fingers are positioned in a semi-circular or C-shaped arrangement.[77] Each zinc finger is made up of anti-parallel beta sheets and an alpha helix, held together by a zinc ion and hydrophobic residues.[76][77]
The zinc atom is constrained in a tetrahedral conformation through the coordination of Cys3, Cys6, His19, and His23 and Zinc – Sulfur bond distance of 2.30 +/- 0.05 Angstroms and Zinc – Nitrogen bond distances of 2.0 +/- 0.05 Angstroms.[77][78][79]
Each zinc finger has an arginine (arg) amino acid protruding from the alpha helix, which forms a hydrogen bond with Nitrogen 7 and Oxygen 6 of the guanine (gua) that is located at the 3’ end of the binding site.[76][77][79] The arg-gua bond is stabilized by aspartic acid from a 2nd residue, which positions the long chain of arginine through a hydrogen bond salt bridge interaction.[76][80]
In residue 3 of the 2nd (i.e., middle) zinc finger, histidine49 forms a hydrogen bond with a co-planar guanine in base pair 6. The stacking of Histidine against Thymine in base pair 5 limits the conformational ability of Histidine49 leading to increased specificity for the histidine-guanine hydrogen bond.[76][77]
At the 6th residue, fingers 1 and 3 have arginine donating a pair of charged hydrogen bonds to Nitrogen 7 and Oxygen 6 of guanine at the 5’ end enhancing the site recognition sequence of zinc fingers.[76][77]
Contacts with DNA backbone
The histidine coordinated to the zinc atom, which is also the seventh residue in the alpha helix of the zinc fingers, coordinates the Zinc ion through its Nε and hydrogen bonds with phosphodiester oxygen through Nδ on the primary DNA strand.[76][77][80]
In addition to histidine, a conserved arginine on the second beta strand of the zinc fingers makes contact with the phosphodiester oxygen on the DNA strand.[76][77][80]
Also serine 75 on the third finger hydrogen bonds to the phosphate between base pairs 7 and 8, as the only backbone contact with the secondary strand of DNA.[76][77][80]
Nuclease dimerization and cleavage
It has been discovered that FokI has no intrinsic specificity in its cleavage of DNA and that the zinc finger recognition domain confers selectivity to zinc finger nucleases.[3][61]
Specificity is provided by dimerization, which decreases the probability of off-site cleavage. Each set of zinc fingers is specific to a nucleotide sequence on either side of the targeted gene 5-7 bp separation between nuclease components.[3]
The dimerization of two ZFNs is required to produce the necessary double-strand break within the CCR5 gene because the interaction between the FokI enzyme and DNA is weak.[60] This break is repaired by the natural repair mechanisms of the cell, specifically non-homologous end joining.[60]
Introducing the CCR5 mutation
Introducing genome alterations depends upon either of the two natural repair mechanisms of a cell: non-homologous end joining (NHEJ) and homology-directed repair (HDR).[60] Repair through NHEJ comes about by the ligation of the end of the broken strands and, upon the occurrence of an error, can produce small insertions and deletions. HDR, on the other hand, makes use of a homologous DNA strand in order to repair and gene and making use of this repair mechanism and providing the desired nucleotide sequence allows for gene insertion or modification.[60]
In the absence of a homologous nucleotide base sequence that can be used by a homologous recombination mechanism, the main DSB repair pathway in mammals is through non-homologous end joining (NHEJ).[81] NHEJ, although capable of restoring a damaged gene, is error-prone.[81] DSB are, therefore, introduced into the gene until an error in its repair occurs at which point ZFNs are no longer able to bind and dimerize and the mutation is complete.[81] In order to accelerate this process, exonucleases can be introduced to digest the ends of the strands generated at DSBs.[81]
Limitations
Increasing the number of zinc fingers increases the specificity by increasing the number of base pairs that the ZFN can bind to.[3] However too many zinc fingers can lead to off-target binding and thus offsite cleavage.[3] This is due to an increased likelihood of zinc fingers binding to parts of the genome outside of the gene of interest.
Current ZFN treatments focus on the CCR5 gene as no known side effects result from altering CCR5.[82] There are strains of HIV that are able to use CXCR4 to enter the host cell, bypassing CCR5 altogether.[82] The same gene editing technology has been applied to CXCR4 alone and in combination with CCR5 [83][84]
Several issues exist with this experimental treatment. One issue lies in ensuring that the desired repair mechanism is the one that is used to repair the DSB following gene addition.[85] Another issue with the disruption of the CCR5 gene is that CXCR4-specific or dual-tropic strains are still able to access the cell.[85] This method can prevent the progression of HIV infection.
To employ the ZFNs in clinical settings the following criteria need to be met:
i) High specificity of DNA-binding – Correlates with better performance and less toxicity of ZFNs. Engineered ZFNs take into account positional and context-dependent effects of zinc fingers to increase specificity.[86]
ii) Enable allosteric activation of FokI once bound to DNA in order for it to produce only the required DSB.[86]
iii) In order to deliver two different zinc finger nuclease subunits and donor DNA to the cell, the vectors that are used need to be improved to decrease the risk of mutagenesis.[86] These include adeno-associated virus vectors, integrase-deficient lentiviral vectors and adenovirus type 5 vectors.[86]
iv) Transient expression of ZFNs would be preferred over permanent expression of these proteins in order to avoid ‘off-target’ effects.[86]
v) During gene targeting, genotoxicity associated with high expression of ZFNs might lead to cell apoptosis and thus needs to be thoroughly verified in vitro and in vivo transformation assays.[86]
Administration of treatment
The cells in which the mutations are induced ex vivo are filtered out from lymphocytes by apheresis to produce analogous lentiviral engineered CD4+ T-cells.[87] These are re-infused into the body as a single dose of 1 X 1010 gene modified analogous CD4+ T-cells.[87] A viral vector is used to deliver the ZFNs that will induce the desired mutation into the cells. Conditions that promote this process are carefully monitored ensuring the production of CCR5 strain HIV-resistant T cells.[88]
Timothy Ray Brown, who underwent a bone marrow transplant in 2007 to treat leukemia, had HIV simultaneously.[89] Soon after the operation the HIV dropped to undetectable levels.[89] This is a result of the bone marrow donor being homozygous for the CCR5-Δ32 mutation.[89] This new mutation conferred a resistance to HIV in the recipient, eventually leading to an almost complete disappearance of HIV particles in his body.[89] After nearly 2 years without antiretroviral drug therapy, HIV could still not be detected in any of his tissues.[89][90] Though this method has been effective at reducing the level of infection, the risks associated with bone marrow transplants outweighs its potential value as a treatment for HIV.[53]
See also
- Genome editing with engineered nucleases
- Zinc finger
- Gene targeting
- Zinc finger protein
- Zinc finger chimera
- Protein engineering
- Genome engineering
References
- ↑ Ramirez CL, Foley JE, Wright DA, et al. (May 2008). "Unexpected failure rates for modular assembly of engineered zinc fingers". Nat. Methods. 5 (5): 374–5. doi:10.1038/nmeth0508-374. PMID 18446154.
- ↑ Maeder ML, et al. (September 2008). "Rapid "open-source" engineering of customized zinc-finger nucleases for highly efficient gene modification". Mol. Cell. 31 (2): 294–301. doi:10.1016/j.molcel.2008.06.016. PMC 2535758
 . PMID 18657511.
. PMID 18657511. - 1 2 3 4 5 6 7 8 9 Carroll, D (2011). "Genome engineering with zinc-finger nucleases". Genetics Society of America. 188 (4): 773–782. doi:10.1534/genetics.111.131433.
- ↑ Kim, YG; Cha, J.; Chandrasegaran, S. (1996). "Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain". Proc Natl Acad Sci USA. 93 (3): 1156–60. Bibcode:1996PNAS...93.1156K. doi:10.1073/pnas.93.3.1156. PMC 40048. PMID 8577732.
- ↑ Bitinaite, J.; D. A. Wah, Aggarwal, A. K., Schildkraut, I. (1998). "FokI dimerization is required for DNA cleavage". Proc Natl Acad Sci USA. 95 (18): 10570–5. Bibcode:1998PNAS...9510570B. doi:10.1073/pnas.95.18.10570. PMC 27935. PMID 9724744.
- ↑ Cathomen T, Joung JK (July 2008). "Zinc-finger nucleases: the next generation emerges". Mol. Ther. 16 (7): 1200–7. doi:10.1038/mt.2008.114. PMID 18545224.
- ↑ Guo, J.; Gaj, T.; Barbas Iii, C. F. (2010). "Directed Evolution of an Enhanced and Highly Efficient FokI Cleavage Domain for Zinc Finger Nucleases". Journal of Molecular Biology. 400 (1): 96–107. doi:10.1016/j.jmb.2010.04.060. PMC 2885538. PMID 20447404.
- ↑ Szczepek, M.; Brondani, V.; Büchel, J.; Serrano, L.; Segal, D. J.; Cathomen, T. (2007). "Structure-based redesign of the dimerization interface reduces the toxicity of zinc-finger nucleases". Nature Biotechnology. 25 (7): 786–793. doi:10.1038/nbt1317. PMID 17603476.
- ↑ Miller, J. C.; Holmes, M. C.; Wang, J.; Guschin, D. Y.; Lee, Y. L.; Rupniewski, I.; Beausejour, C. M.; Waite, A. J.; Wang, N. S.; Kim, K. A.; Gregory, P. D.; Pabo, C. O.; Rebar, E. J. (2007). "An improved zinc-finger nuclease architecture for highly specific genome editing". Nature Biotechnology. 25 (7): 778–785. doi:10.1038/nbt1319. PMID 17603475.
- ↑ Doyon, Y.; Vo, T. D.; Mendel, M. C.; Greenberg, S. G.; Wang, J.; Xia, D. F.; Miller, J. C.; Urnov, F. D.; Gregory, P. D.; Holmes, M. C. (2010). "Enhancing zinc-finger-nuclease activity with improved obligate heterodimeric architectures". Nature Methods. 8 (1): 74–79. doi:10.1038/nmeth.1539. PMID 21131970.
- ↑ Ramalingam, S.; Kandavelou, K.; Rajenderan, R.; Chandrasegaran, S. (2011). "Creating Designed Zinc-Finger Nucleases with Minimal Cytotoxicity". Journal of Molecular Biology. 405 (3): 630–641. doi:10.1016/j.jmb.2010.10.043. PMC 3017627. PMID 21094162.
- ↑ Zhang, F.; Maeder, M. L.; Unger-Wallace, E.; Hoshaw, J. P.; Reyon, D.; Christian, M.; Li, X.; Pierick, C. J.; Dobbs, D.; Peterson, T.; Joung, J. K.; Voytas, D. F. (2010). "High frequency targeted mutagenesis in Arabidopsis thaliana using zinc finger nucleases". Proceedings of the National Academy of Sciences. 107 (26): 12028–12033. doi:10.1073/pnas.0914991107.
- ↑ Osakabe, K.; Osakabe, Y.; Toki, S. (2010). "Site-directed mutagenesis in Arabidopsis using custom-designed zinc finger nucleases". Proceedings of the National Academy of Sciences. 107 (26): 12034–12039. doi:10.1073/pnas.1000234107.
- ↑ Cai, C. Q.; Doyon, Y.; Ainley, W. M.; Miller, J. C.; Dekelver, R. C.; Moehle, E. A.; Rock, J. M.; Lee, Y. L.; Garrison, R.; Schulenberg, L.; Blue, R.; Worden, A.; Baker, L.; Faraji, F.; Zhang, L.; Holmes, M. C.; Rebar, E. J.; Collingwood, T. N.; Rubin-Wilson, B.; Gregory, P. D.; Urnov, F. D.; Petolino, J. F. (2008). "Targeted transgene integration in plant cells using designed zinc finger nucleases". Plant Molecular Biology. 69 (6): 699–709. doi:10.1007/s11103-008-9449-7. ISSN 0167-4412. PMID 19112554.
- ↑ Townsend, J. A.; Wright, D. A.; Winfrey, R. J.; Fu, F.; Maeder, M. L.; Joung, J. K.; Voytas, D. F. (2009). "High-frequency modification of plant genes using engineered zinc-finger nucleases". Nature. 459 (7245): 442–445. Bibcode:2009Natur.459..442T. doi:10.1038/nature07845. PMC 2743854. PMID 19404258.
- ↑ Curtin, S. J.; Zhang, F.; Sander, J. D.; Haun, W. J.; Starker, C.; Baltes, N. J.; Reyon, D.; Dahlborg, E. J.; Goodwin, M. J.; Coffman, A. P.; Dobbs, D.; Joung, J. K.; Voytas, D. F.; Stupar, R. M. (2011). "Targeted Mutagenesis of Duplicated Genes in Soybean with Zinc-Finger Nucleases". Plant Physiology. 156 (2): 466–473. doi:10.1104/pp.111.172981. PMC 3177250. PMID 21464476.
- ↑ Shukla VK, Doyon Y, Miller JC, et al. (May 2009). "Precise genome modification in the crop species Zea mays using zinc-finger nucleases". Nature. 459 (7245): 437–41. Bibcode:2009Natur.459..437S. doi:10.1038/nature07992. PMID 19404259.
- ↑ Bibikova, M.; Beumer, K.; Trautman, J.; Carroll, D. (2003). "Enhancing Gene Targeting with Designed Zinc Finger Nucleases". Science. 300 (5620): 764. doi:10.1126/science.1079512. PMID 12730594.
- 1 2 Wood, A. J.; Lo, T. -W.; Zeitler, B.; Pickle, C. S.; Ralston, E. J.; Lee, A. H.; Amora, R.; Miller, J. C.; Leung, E.; Meng, X.; Zhang, L.; Rebar, E. J.; Gregory, P. D.; Urnov, F. D.; Meyer, B. J. (2011). "Targeted Genome Editing Across Species Using ZFNs and TALENs". Science. 333 (6040): 307. doi:10.1126/science.1207773. PMC 3489282. PMID 21700836.
- ↑ Gühmann, Martin; Jia, Huiyong; Randel, Nadine; Verasztó, Csaba; Bezares-Calderón, Luis A.; Michiels, Nico K.; Yokoyama, Shozo; Jékely, Gáspár (August 2015). "Spectral Tuning of Phototaxis by a Go-Opsin in the Rhabdomeric Eyes of Platynereis". Current Biology. 25 (17): 2265–2271. doi:10.1016/j.cub.2015.07.017. PMID 26255845.
- ↑ Ochiai, H.; Fujita, K.; Suzuki, K. I.; Nishikawa, M.; Shibata, T.; Sakamoto, N.; Yamamoto, T. (2010). "Targeted mutagenesis in the sea urchin embryo using zinc-finger nucleases". Genes to Cells. 15 (8): no. doi:10.1111/j.1365-2443.2010.01425.x. PMID 20604805.
- ↑ Takasu, Y.; Kobayashi, I.; Beumer, K.; Uchino, K.; Sezutsu, H.; Sajwan, S.; Carroll, D.; Tamura, T.; Zurovec, M. (2010). "Targeted mutagenesis in the silkworm Bombyx mori using zinc finger nuclease mRNA injection". Insect Biochemistry and Molecular Biology. 40 (10): 759–765. doi:10.1016/j.ibmb.2010.07.012. PMID 20692340.
- ↑ S.C. Ekker (2008). "Zinc Finger–Based Knockout Punches for Zebrafish Genes". Zebrafish. 5 (2): 1121–3. doi:10.1089/zeb.2008.9988. PMC 2849655. PMID 18554175.
- ↑ Young, J. J.; Cherone, J. M.; Doyon, Y.; Ankoudinova, I.; Faraji, F. M.; Lee, A. H.; Ngo, C.; Guschin, D. Y.; Paschon, D. E.; Miller, J. C.; Zhang, L.; Rebar, E. J.; Gregory, P. D.; Urnov, F. D.; Harland, R. M.; Zeitler, B. (2011). "Efficient targeted gene disruption in the soma and germ line of the frog Xenopus tropicalis using engineered zinc-finger nucleases". Proceedings of the National Academy of Sciences. 108 (17): 7052–7057. doi:10.1073/pnas.1102030108.
- ↑ Goldberg, A. D.; Banaszynski, L. A.; Noh, K. M.; Lewis, P. W.; Elsaesser, S. J.; Stadler, S.; Dewell, S.; Law, M.; Guo, X.; Li, X.; Wen, D.; Chapgier, A.; Dekelver, R. C.; Miller, J. C.; Lee, Y. L.; Boydston, E. A.; Holmes, M. C.; Gregory, P. D.; Greally, J. M.; Rafii, S.; Yang, C.; Scambler, P. J.; Garrick, D.; Gibbons, R. J.; Higgs, D. R.; Cristea, I. M.; Urnov, F. D.; Zheng, D.; Allis, C. D. (2010). "Distinct Factors Control Histone Variant H3.3 Localization at Specific Genomic Regions". Cell. 140 (5): 678–691. doi:10.1016/j.cell.2010.01.003. PMC 2885838. PMID 20211137.
- ↑ Geurts, A. M.; Cost, G. J.; Freyvert, Y.; Zeitler, B.; Miller, J. C.; Choi, V. M.; Jenkins, S. S.; Wood, A.; Cui, X.; Meng, X.; Vincent, A.; Lam, S.; Michalkiewicz, M.; Schilling, R.; Foeckler, J.; Kalloway, S.; Weiler, H.; Menoret, S.; Anegon, I.; Davis, G. D.; Zhang, L.; Rebar, E. J.; Gregory, P. D.; Urnov, F. D.; Jacob, H. J.; Buelow, R. (2009). "Knockout Rats via Embryo Microinjection of Zinc-Finger Nucleases". Science. 325 (5939): 433–433. doi:10.1126/science.1172447. PMC 2831805. PMID 19628861.
- ↑ Flisikowska, T.; Thorey, I. S.; Offner, S.; Ros, F.; Lifke, V.; Zeitler, B.; Rottmann, O.; Vincent, A.; Zhang, L.; Jenkins, S.; Niersbach, H.; Kind, A. J.; Gregory, P. D.; Schnieke, A. E.; Platzer, J. (2011). Milstone, David S., ed. "Efficient Immunoglobulin Gene Disruption and Targeted Replacement in Rabbit Using Zinc Finger Nucleases". PLoS ONE. 6 (6): e21045. doi:10.1371/journal.pone.0021045. PMC 3113902. PMID 21695153.
- ↑ Hauschild, J.; Petersen, B.; Santiago, Y.; Queisser, A. -L.; Carnwath, J. W.; Lucas-Hahn, A.; Zhang, L.; Meng, X.; Gregory, P. D.; Schwinzer, R.; Cost, G. J.; Niemann, H. (2011). "Efficient generation of a biallelic knockout in pigs using zinc-finger nucleases". Proceedings of the National Academy of Sciences. 108 (29): 12013–12017. doi:10.1073/pnas.1106422108.
- ↑ Yu, S.; Luo, J.; Song, Z.; Ding, F.; Dai, Y.; Li, N. (2011). "Highly efficient modification of beta-lactoglobulin (BLG) gene via zinc-finger nucleases in cattle". Cell Research. 21: 1638–1640. doi:10.1038/cr.2011.153.
- ↑ D. Carroll (2008). "Zinc-finger Nucleases as Gene Therapy Agents". Gene Therapy. 15 (22): 1463–1468. doi:10.1038/gt.2008.145. PMC 2747807. PMID 18784746.
- ↑ Li, H.; Haurigot, V.; Doyon, Y.; Li, T.; Wong, S. Y.; Bhagwat, A. S.; Malani, N.; Anguela, X. M.; Sharma, R.; Ivanciu, L.; Murphy, S. L.; Finn, J. D.; Khazi, F. R.; Zhou, S.; Paschon, D. E.; Rebar, E. J.; Bushman, F. D.; Gregory, P. D.; Holmes, M. C.; High, K. A. (2011). "In vivo genome editing restores haemostasis in a mouse model of haemophilia". Nature. 475 (7355): 217–221. doi:10.1038/nature10177. PMC 3152293. PMID 21706032.
- ↑ Tebas, Pablo; Stein, David; Tang, Winson W.; Frank, Ian; Wang, Shelley Q.; Lee, Gary; Spratt, S. Kaye; Surosky, Richard T.; Giedlin, Martin A.; Nichol, Geoff; Holmes, Michael C.; Gregory, Philip D.; Ando, Dale G.; Kalos, Michael; Collman, Ronald G.; Binder-Scholl, Gwendolyn; Plesa, Gabriela; Hwang, Wei-Ting; Levine, Bruce L.; June, Carl H. (6 March 2014). "Gene Editing of CCR5 in Autologous CD4 T Cells of Persons Infected with HIV". New England Journal of Medicine. 370 (10): 901–910. doi:10.1056/NEJMoa1300662. PMC 4084652. PMID 24597865.
- 1 2 3 4 Durai S, Mani M, Kandavelou K, Wu J, Porteus MH, Chandrasegaran S (2005). "Zinc finger nucleases: custom-designed molecular scissors for genome engineering of plant and mammalian cells". Nucleic Acids Res. 33 (18): 5978–90. doi:10.1093/nar/gki912. PMC 1270952. PMID 16251401.
- ↑ Lee HJ, Kim E, Kim JS (December 2009). "Targeted chromosomal deletions in human cells using zinc finger nucleases". Genome Res. 20 (1): 81–9. doi:10.1101/gr.099747.109. PMC 2798833. PMID 19952142.
- ↑ Mittelman, D; Moye, C; Morton, J; Sykoudis, K; Lin, Y; Carroll, D; Wilson, JH (2009-06-16). "Zinc-finger directed double-strand breaks within CAG repeat tracts promote repeat instability in human cells". Proceedings of the National Academy of Sciences of the United States of America. 106 (24): 9607–12. Bibcode:2009PNAS..106.9607M. doi:10.1073/pnas.0902420106. PMC 2701052. PMID 19482946.
- ↑ Kandavelou K, Chandrasegaran S (2008). "Plasmids for Gene Therapy". Plasmids: Current Research and Future Trends. Caister Academic Press. ISBN 978-1-904455-35-6.
- ↑ Gupta A, Meng X, Zhu LJ, Lawson ND, Wolfe SA (September 2010). "Zinc finger protein-dependent and -independent contributions to the in vivo off-target activity of zinc finger nucleases". Nucleic Acids Res. 39 (1): 381–392. doi:10.1093/nar/gkq787. PMC 3017618. PMID 20843781..
- ↑ Pattanayak, V.; Ramirez, C. L.; Joung, J. K.; Liu, D. R. (2011). "Revealing Off-Target Cleavage Specificities of Zinc Finger Nucleases by in Vitro Selection". Nature Methods. 8 (9): 765–770. doi:10.1038/nmeth.1670. PMC 3164905. PMID 21822273.
- 1 2 Liu, X.; Wang, Y.S; Guo, W.J.; Chang, B.H.; Liu, J.; Guo, Z.K.; Quan, F.S.; Zhang, Y. (2013). "Zinc-finger nickase-mediated insertion of the lysostaphin gene into the beta-casein locus in cloned cows". Nature Communication. 4. doi:10.1038/ncomms3565.
- ↑ Liu, X.; Wang, Y.; Tian, Y.; Yu, Y.; Gao, M.; Hu, G.; Su, F.; Pan, S.; Luo, Y.; Guo, Z.; Quan, F.; Zhang, Y. (2014). "Generation of mastitis resistance in cows by targeting human lysozyme gene to -casein locus using zinc-finger nucleases". Proceedings of the Royal Society B: Biological Sciences. 281 (1780): 20133368. doi:10.1098/rspb.2013.3368.
- ↑ Grizot S, Smith J, Daboussi F, et al. (September 2009). "Efficient targeting of a SCID gene by an engineered single-chain homing endonuclease". Nucleic Acids Res. 37 (16): 5405–19. doi:10.1093/nar/gkp548. PMC 2760784. PMID 19584299.
- ↑ Gao, H.; Smith, J.; Yang, M.; Jones, S.; Djukanovic, V.; Nicholson, M. G.; West, A.; Bidney, D.; Falco, S. C.; Jantz, D.; Lyznik, L. A. (2010). "Heritable targeted mutagenesis in maize using a designed endonuclease". The Plant Journal. 61 (1): 176–187. doi:10.1111/j.1365-313X.2009.04041.x. PMID 19811621.
- ↑ Christian M, Cermak T, Doyle EL, et al. (July 2010). "Targeting DNA Double-Strand Breaks with TAL Effector Nucleases". Genetics. 186 (2): 757–61. doi:10.1534/genetics.110.120717. PMC 2942870. PMID 20660643.
- ↑ Li T, Huang S, Jiang WZ, et al. (August 2010). "TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain". Nucleic Acids Res. 39 (1): 359–372. doi:10.1093/nar/gkq704. PMC 3017587. PMID 20699274.
- ↑ Moscou MJ, Bogdanove AJ; Bogdanove (December 2009). "A simple cipher governs DNA recognition by TAL effectors". Science. 326 (5959): 1501. Bibcode:2009Sci...326.1501M. doi:10.1126/science.1178817. PMID 19933106.
- ↑ Boch J, Scholze H, Schornack S, et al. (December 2009). "Breaking the code of DNA binding specificity of TAL-type III effectors". Science. 326 (5959): 1509–12. Bibcode:2009Sci...326.1509B. doi:10.1126/science.1178811. PMID 19933107.
- ↑ Miller, J. C.; Tan, S.; Qiao, G.; Barlow, K. A.; Wang, J.; Xia, D. F.; Meng, X.; Paschon, D. E.; Leung, E.; Hinkley, S. J.; Dulay, G. P.; Hua, K. L.; Ankoudinova, I.; Cost, G. J.; Urnov, F. D.; Zhang, H. S.; Holmes, M. C.; Zhang, L.; Gregory, P. D.; Rebar, E. J. (2010). "A TALE nuclease architecture for efficient genome editing". Nature Biotechnology. 29 (2): 143–148. doi:10.1038/nbt.1755. PMID 21179091.
- ↑ Wade, Nicholas (28 December 2009). "Zinc Fingers Could Be Key to Reviving Gene Therapy". The New York Times. New York Times. Retrieved 31 May 2016.
- 1 2 Ramirez, C. L.; Certo, M. T.; Mussolino, C.; Goodwin, M. J.; Cradick, T. J.; McCaffrey, A. P.; Cathomen, T.; Scharenberg, A. M.; Joung, J. K. (2012). "Engineered zinc finger nickases induce homology-directed repair with reduced mutagenic effects". Nucleic Acids Research. 40 (7): 5560–5568. doi:10.1093/nar/gks179.
- ↑ Wang, J.; Friedman, G.; Doyon, Y.; Wang, N. S.; Li, C. J.; Miller, J. C.; Hua, K. L.; Yan, J. E.; Babiarz, P. D.; Gregory, P. D.; Holmes, M. C. (2012). "Targeted gene addition to a predetermined site in the human genome using a ZFN-based nicking enzyme". Genome Research. 22 (4): 1316–1326. doi:10.1101/gr.122879.111.
- ↑ Deeks, S. G.; McCune, J. M. (2010). "Can HIV be cured with stem cell therapy?.". Nature Biotechnology. 28 (8): 807–810. doi:10.1038/nbt0810-807.
- ↑ Alkhatib, G (2009). "The biology of CCR5 and CXCR4". Current Opinion in HIV and AIDS. 4 (2): 96–103. doi:10.1097/coh.0b013e328324bbec.
- 1 2 Hütter, G.; Nowak, D.; Mossner, M.; Ganepola, S.; Müßig, A.; Allers, K.; Thiel, E. (2009). "Long-term control of HIV by CCR5 Delta32/Delta32 stem-cell transplantation". New England Journal of Medicine. 360 (7): 692–698. doi:10.1056/nejmoa0802905.
- ↑ Carroll, D (2008). "Progress and prospects: zinc-finger nucleases as gene therapy agents". Gene Therapy. 15 (22): 1463–1468. doi:10.1038/gt.2008.145. PMC 2747807. PMID 18784746.
- ↑ Perez, E. E.; Wang, J.; Miller, J. C.; Jouvenot, Y.; Kim, K. A.; Liu, O.; June, C.H. (2008). "Establishment of HIV-1 resistance in CD4+ T cells by genome editing using zinc-finger nucleases". Nature Biotechnology. 26 (7): 808–816. doi:10.1038/nbt1410.
- ↑ Chung, J.; Rossi, J. J.; Jung, U. (2011). "Current progress and challenges in HIV gene therapy". Future virology. 6 (11): 1319–1328. doi:10.2217/fvl.11.113.
- 1 2 Lai, Y. CCR5-targeted hematopoietic stem cell gene approaches for HIV disease: Current progress and future prospects Current Stem Cell Research and Therapy, 2012; 7 (4), pp. 310-317.
- ↑ De Silva, E., Stumpf, Michael P.H. (2004). "HIV and the CCR5-D32 Resistance Allele.". FEMS Microbiology Letters. 241 (1): 1–12. doi:10.1016/j.femsle.2004.09.040. PMID 15556703.
- ↑ Durand, Christine. M, Siliciano, Robert F. (2014). "Dual Zinc-Finger Nucleases Block HIV Infection.". Blood Journal. 123 (1): 636–646.
- 1 2 3 4 5 Urnov, F. D.; Rebar, E. J.; Holmes, M. C.; Zhang, H. S.; Gregory, P. D. (2010). "Genome editing with engineered zinc finger nucleases". Nature Reviews Genetics. 11 (9): 636–646. doi:10.1038/nrg2842. PMID 20717154.
- 1 2 Urnov, F. D.; Miller, J. C.; Lee; Beausejour; Rock, J. M.; Augustus, S.; Holmes, M. C. (2005). "Highly efficient endogenous human gene correction using designed zinc-finger nucleases". Nature. 435 (7042): 646–651. doi:10.1038/nature03556. PMID 15806097.
- ↑ Wayengera, M. "Proviral HIV-genome-wide and pol-gene specific zinc finger nucleases: usability for targeted HIV gene therapy. Theor Biol Med Model, 2011; 8, pp26.
- ↑ Wayengera, M.Zinc finger arrays binding human papillomavirus types 16 and 18 genomic DNA: precursors of gene-therapeutics for in-situ reversal of associated cervical neoplasia.Theor Biol Med Model, (2011), 9, pp30.
- ↑ Wayengera, M.Identity of zinc finger nucleases with specificity to herpes simplex virus type II genomic DNA: novel HSV-2 vaccine/therapy precursors.Theor Biol Med Model, (2011), 8, pp23.
- ↑ Wayengera, M (2003). "HIV and Gene Therapy: The proposed [R-M enzymatic] model for a gene therapy against HIV". Makerere Med J. 38: 28–30.
- ↑ Wayengera, M; Kajumbula, H; Byarugaba, W (2007). "Frequency and site mapping of HIV-1/SIVcpz, HIV-2/SIVsmm and Other SIV gene sequence cleavage by various bacteria restriction enzymes: Precursors for a novel HIV inhibitory product". Afr J Biotechnol. 6 (10): 1225–1232.
- ↑ Wayengera M, Kajumbula H, Byarugaba W: Identification of restriction endonuclease with potential ability to cleave the HSV-2 genome: inherent potential for biosynthetic versus live microbicides. Theor Biol Med Model. 2008, 5:18.
- ↑ Wayengera, M (2008). "Pre-Integration gene slicing (PRINT-GSX) as an alternate or complimentary gene therapy modem to RNA interference". J Appl Biol Sci. 1 (2): 56–63.
- ↑ Wayengera M: Diverting primary HIV entry and replication to vaginal commensal lactobacillus expressing R-M nucleic enzymatic peptides with potent activity at cleaving proviral DNA as a novel HIV live microbicide strategy. Microbicide- New Delhi, India 2008. Abs-10.
- ↑ Wayengera M: Preparing for a Phase 1 Preclinical trial of VRX-SMR: a Lentiviral Vector transduced with restriction enzymes cleaving HIV proviral DNA as a therapeutic vaccine: Opportunities and Challenges. Vaccine Congress -Amsterdam, Netherlands 2007, :24OR.
- ↑ Wayengera M: xREPLAB: A recombinant lactobacillus strain producing restriction enzymes with potent activity against HIV proviral DNA as a Live Microbicide Strategy. AIDS vaccine- Washington, Seattle 2007, :P05-01.
- ↑ Wayengera, M (2007). "A Recombinant lactobacillus strain producing restriction enzymes that cleave proviral HIV DNA may offer a novel Vagino-cervical mucosal exogenous live Microbicide strategy for preventing HIV transmission among high-risk women". Afr J Biotechnol. 6 (15): 1750–1756.
- ↑ Wayengera, M (2007). "PREX-1979: Modeling the first ever prototype of could be a 5th generation of Microbicides for preventing HIV infection among high-risk women". Afr J Biotechnol. 6 (10): 1221–1224.
- ↑ Wayengera, M (2008). "Why bacteria derived R-M nucleic enzymatic peptides are efficient therapeutic molecules for use in the design and development of novel HIV therapeutic molecules". Afr J Biotechnol. 7 (12): 1791–1796.
- 1 2 Urnov, F. D., Rebar, E. J., Holmes, M. C., Zhang, H. S., & Gregory, P. D. (2010). "Genome Editing with Engineered Zinc Finger Nucleases.". Nature Reviews.Genetics. 11 (9): 636–646. doi:10.1038/nrg2842. PMID 20717154.
- 1 2 3 4 5 6 7 8 9 10 11 Pavletich, N. P.; Pabo, C. O. (1991). "Zinc finger-DNA recognition: crystal structure of a Zif268-DNA complex at 2.1 A.". Science. 252 (5007): 809–817. doi:10.1126/science.2028256. PMID 2028256.
- 1 2 3 4 5 6 7 8 9 10 11 Klug, A (2005). "The discovery of zinc fingers and their development for practical applications in gene regulation". Proceedings of the Japan Academy, Series B. 81 (4): 87–102. doi:10.2183/pjab.81.87.
- ↑ Frankel, A. D.; Berg, J. M.; Pabo, C. O. (1987). "Metal-dependent folding of a single zinc finger from transcription factor IIIA". Proceedings of the National Academy of Sciences. 84 (14): 4841–4845. doi:10.1073/pnas.84.14.4841.
- 1 2 Lee, M. S.; Gippert, G. P.; Soman, K. V.; Case, D. A.; Wright, P. E. (1989). "Three-dimensional solution structure of a single zinc finger DNA-binding domain". Science. 245 (4918): 635–637. doi:10.1126/science.2503871.
- 1 2 3 4 Klug, A.; Schwabe, J. W. (1995). "Protein motifs 5. Zinc fingers". The FASEB Journal. 9 (8): 597–604.
- 1 2 3 4 Stone, D.; Kiem, H. P.; & Jerome, K. R. Targeted gene disruption to cure HIV. Curr Opin HIV AIDS. 2013, 8, pp 000-000.
- 1 2 Coakley, E., Petropoulos, C.J., Whitcomb, J.M. Assessing ch vbgemokine co-receptor usage in HIV. Curr. Opin. Infect. Dis. (2005) 18(1), pp 9-15.
- ↑ Wilen, C.B.; Wang, J.; Tilton, J.C.; et al. (2011). "Engineering HIV-resistant humant CD4+ T cells with CXCR4-specific zinc-finger nucleases". PLoS Pathogens.
- ↑ Didigu, C.A.; Wilen, C.B.; Wang, J. (2013). "Simultaneous zinc-finger nuclease editing of the HIV coreceptors ccr5 and cxcr4 protects CD4+ T cells from HIV-1 infection". Blood.
- 1 2 Barton, K. M.; Burch, B. D.; Soriano-Sarabia, N.; & Margolis, D. M. Prospects for treatment of latent HIV. Clinical Pharmacology & Therapeutics. 2012.
- 1 2 3 4 5 6 Cathomen, T., & Joung, J. K.. Zinc-finger nucleases: the next generation emerges. Molecular Therapy, (2008) 16(7), pp 1200-1207.
- 1 2 Levine, B. L.; Humeau, L. M.; Boyer, J.; MacGregor, R. R.; Rebello, T.; Lu, X.; June, C. H. (2006). "Gene transfer in humans using a conditionally replicating lentiviral vector". Proceedings of the National Academy of Sciences. 103 (46): 17372–17377. doi:10.1073/pnas.0608138103. PMC 1635018. PMID 17090675.
- ↑ Varela-Rohena, A.; Carpenito, C.; Perez, E. E.; Richardson, M.; Parry, R. V.; Milone, M.; Riley, J. L. (2008). "Genetic engineering of T cells for adoptive immunotherapy". Immunologic research. 42 (1-3): 166–181. doi:10.1007/s12026-008-8057-6. PMC 2699549. PMID 18841331.
- 1 2 3 4 5 Rosenberg, T. "The man who had HIV and now does not". New York Magazine. Retrieved January 2013.
- ↑ Hütter G, Ganepola S (2011). "Eradication of HIV by transplantation of CCR5-deficient hematopoietic stem cells". The Scientific World Journal. 11: 1068–1076. doi:10.1100/tsw.2011.102. PMID 21552772.
Further reading
- Mandell JG, Barbas CF (July 2006). "Zinc Finger Tools: custom DNA-binding domains for transcription factors and nucleases". Nucleic Acids Res. 34 (Web Server issue): W516–23. doi:10.1093/nar/gkl209. PMC 1538883. PMID 16845061.
- Porteus MH, Carroll D (August 2005). "Gene targeting using zinc finger nucleases". Nat. Biotechnol. 23 (8): 967–73. doi:10.1038/nbt1125. PMID 16082368.
- Doyon Y, McCammon JM, Miller JC, et al. (June 2008). "Heritable Targeted Gene Disruption in Zebrafish Using Designed Zinc Finger Nucleases". Nat. Biotechnol. 26 (6): 702–8. doi:10.1038/nbt1409. PMC 2674762. PMID 18500334.
- Meng X, Noyes MB, Zhu LJ, Lawson ND, Wolfe SA (June 2008). "Targeted gene inactivation in zebrafish using engineered zinc finger nucleases". Nat. Biotechnol. 26 (6): 695–701. doi:10.1038/nbt1398. PMC 2502069. PMID 18500337.
External links
- Zinc finger selector
- Zinc Finger Consortium website
- Zinc Finger Consortium materials from Addgene
- A commercial supplier of ZFNs