Aequorin
| Aequorin 1 | |
|---|---|
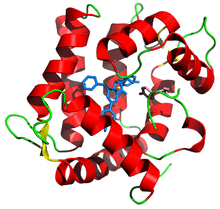 Aequorin ribbon diagram from PDB 1ej3 with prosthetic group coelenterazine in blue | |
| Identifiers | |
| Organism | |
| Symbol | N/A |
| UniProt | P07164 |
| Other data | |
| EC number | 1.13.12.5 |
Aequorin is a calcium-activated photoprotein isolated from the hydrozoan Aequorea victoria.[1] Though the bioluminescence was studied decades before, the protein was originally isolated from the animal by Osamu Shimomura.[2] In the animals, the protein occurs together with the Green fluorescent protein to produce green light by resonant energy transfer, while aequorin by itself generates blue light.
Discussions of "jellyfish DNA" to make "glowing" animals often refer to transgenic animals which express the Green fluorescent protein, not aequorin, although both originally derived from the same animal.
Discovery
Work on aequorin began with E. Newton Harvey in 1921.[3] Though Harvey was unable to demonstrate a classical luciferase-luciferin reaction, he showed that water could produce light from dried photocytes and that light could be produced even in the absence of oxygen. Later, Osamu Shimomura began work into the bioluminescence of Aequorea in 1961. This involved tedious harvesting of tens of thousands of jellyfish from the docks in Friday Harbor, Washington.[1] It was determined that light could be produced from extracts with seawater, and more specifically, with calcium.[2] It was also noted during the extraction the animal creates green light due to the presence of the green fluorescent protein, which changes the native blue light of aequorin to green.[4]
While the main focus of his work was on the bioluminescence,[5] Shimomura and two others, Martin Chalfie and Roger Tsien were awarded the Nobel Prize in 2008 for their work on green fluorescent proteins.
Structure
Aequorin is a holoprotein composed of two distinct units, the apoprotein that is called apoaequorin, which has an approximate molecular weight of 21 kDa, and the prosthetic group coelenterazine, the luciferin.[6] This is to say, apoaequorin is the enzyme produced in the photocytes of the animal, and coelenterazine is the substrate whose oxidation the enzyme catalyzes. When coelenterazine is bound, it is called aequorin. Notably, the protein contains three EF hand motifs that function as binding sites for Ca2+ ions.[7] The protein is a member of the superfamily of the calcium-binding proteins of which there are some 66 subfamilies.[8]
The crystal structure revealed that aequorin binds coelenterazine and oxygen in the form of a peroxide, coelenterazine-2-hydroperoxide.[9] The binding site for the first two calcium atoms show a 20X greater affinity for calcium than the third site.[10] However, earlier claims that only two EF-hands bind calcium,[11] were questioned when later structures indicated that all three site indeed can bind calcium.[12] Thus, titration studies show that all three calcium-binding sites are active but only two ions are needed to trigger the enzymatic reaction.[13]
Other studies have shown the presence of an internal cysteine bond that maintains the structure of aequorin.[14] This has also explained the need for a thiol reagent like beta mercaptoethanol in the regeneration of the protein since such reagents weaken the sulfhydryl bonds between cysteine residues, expediting the regeneration of the aequorin.
Chemical characterization of aequorin indicates the protein is somewhat resilient to harsh treatments. Aequorin is heat resistant.[15] Held at 95⁰C for 2 minutes the protein lost only 25% activity. Denaturants 6M-urea or 4M-guanidine hydrochloride did not destroy the protein.
Genetics
Aequorin is presumably encoded in the genome of Aequorea. At least four copies of the gene were recovered as cDNA from the animal.[16][17] Because the genome has not been sequenced, it is unclear if the cDNA variants can account for all of the isoforms of the protein.[18]
Mechanism of action
Early studies of the bioluminescence of Aequorea by E. Newton Harvey had noted that the bioluminescence appears as a ring around the bell, and occurs even in the absence of air.[19] This was remarkable because most bioluminescence reactions appeared to require oxygen, and led to the idea that the animals somehow store oxygen.[20] It was later discovered that the apoprotein can stably bind coelenterazine and oxygen is required for the regeneration to the active form of aequorin.[21] However, in the presence of calcium ions, the protein undergoes a conformational change and through oxidation converts its prosthetic group, coelenterazine, into excited coelenteramide and CO2.[22] As the excited coelenteramide relaxes to the ground state, blue light (wavelength of 465 nm) is emitted. Before coelenteramide is exchanged out, the entire protein is still fluorescent blue.[23][24] Because of the connection between bioluminescence and fluorescence, this property was ultimately important in the discovery of the luciferin coelenterazine.[25]
Applications
Since the emitted light can be easily detected with a luminometer, aequorin has become a useful tool in molecular biology for the measurement of intracellular Ca2+ levels.[26] The early successful purification of aequorin led to the first experiments involving the injection of the protein into the tissues of living animals to visualize the physiological release of calcium in the muscle fibers of a barnacle.[27] Since then, the protein has been widely used as reported in many model biological systems, including zebrafish,[28] rats, mice, and cultured cells.[29][30][31][32]
Cultured cells expressing the aequorin gene can effectively synthesize apoaequorin: however, recombinant expression yields only the apoprotein, therefore it is necessary to add coelenterazine into the culture medium of the cells to obtain a functional protein and thus use its blue light emission to measure Ca2+ concentration. Coelenterazine is a hydrophobic molecule, and therefore is easily taken up across plant and fungal cell walls, as well as the plasma membrane of higher eukaryotes, making aequorin suitable as a (Ca2+ reporter) in plants, fungi, and mammalian cells.[33][34]
Aequorin has a number of advantages over other Ca2+ indicators: because the protein is large, it has a low leakage rate from cells compared to lipophilic dyes such as DiI. It lacks phenomena of intracellular compartmentalization or sequestration as is often seen for Voltage-sensitive dyes, and does not disrupt cell functions or embryo development. Moreover, the light emitted by the oxidation of coelenterazine does not depend on any optical excitation, so problems with auto-fluorescence are eliminated.[35] The primary limitation of aequorin is that the prosthetic group coelenterazine is irreversibly consumed to produce light, and requires continuous addition of coelenterazine into the media. Such issues led to developments of other genetically encoded calcium sensors including the calmodulin-based sensor cameleon,[36] developed by Roger Tsien and the troponin-based sensor, TN-XXL, developed by Oliver Griesbeck.[37]
See also
- Obelin, a similar photoprotein from Obelia
References
- 1 2 Shimomura O (1995). "A short story of aequorin.". Biol Bull. Biological Bulletin, Vol. 189, No. 1. 189 (1): 1–5. doi:10.2307/1542194. JSTOR 1542194. PMID 7654844.
- 1 2 Shimomura O, Johnson FH, Saiga Y (1962). "Extraction, purification and properties of aequorin, a bioluminescent protein from the luminous hydromedusan, Aequorea". J Cell Comp Physiol. 59: 223–39. doi:10.1002/jcp.1030590302. PMID 13911999.
- ↑ Harvey EN (1921). "Studies on Bioluminescence. XIII. Luminescence in the Cœlenterate". Biological Bulletin. 41 (5): 280–287. doi:10.2307/1536528. JSTOR 1536528.
- ↑ Morin JG, Hastings JW (1971). "Energy transfer in a bioluminescent system". J. Cell. Physiol. 77 (3): 313–8. doi:10.1002/jcp.1040770305. PMID 4397528.
- ↑ Shimomura O (2005). "The discovery of aequorin and green fluorescent protein". J Microsc. 217 (Pt 1): 1–15. doi:10.1111/j.0022-2720.2005.01441.x. PMID 15655058.
- ↑ Shimomura O, Johnson FH (1978). "Peroxidized coelenterazine, the active group in the photoprotein aequorin". PNAS USA. 75 (3): 2611–2615. doi:10.1073/pnas.75.6.2611. PMC 392612
 . PMID 275832.
. PMID 275832. - ↑ Charbonneau H, Walsh KA, McCann RO, Prendergast FG, Cormier MJ, Vanaman TC (1985). "Amino acid sequence of the calcium-dependent photoprotein aequorin". Biochemistry. 24 (24): 6762–71. doi:10.1021/bi00345a006. PMID 2866797.
- ↑ Zhou Y, Yang W, Kirberger M, Lee HW, Ayalasomayajula G, Yang JJ (2006). "Prediction of EF-hand calcium-binding proteins and analysis of bacterial EF-hand proteins". Proteins. 65 (3): 643–55. doi:10.1002/prot.21139. PMID 16981205.
- ↑ Head JF, Inouye S, Teranishi K, Shimomura O (2000). "The crystal structure of the photoprotein aequorin at 2.3 Å resolution". Nature. 405 (6784): 372–6. doi:10.1038/35012659. PMID 10830969.
- ↑ Shimomura O (1995). "Luminescence of aequorin is triggered by the binding of two calcium ions". Biochem. Biophys. Res. Commun. 211 (2): 359–63. doi:10.1006/bbrc.1995.1821. PMID 7794244.
- ↑ Shimomura O (1995). "Luminescence of aequorin is triggered by the binding of two calcium ions". Biochemical and Biophysical Research Communications. 211 (2): 359–363. doi:10.1006/bbrc.1995.1821. PMID 7794244.
- ↑ Deng L, Vysotski ES, Markova SV, Liu ZJ, Lee J, Rose J, Wang BC (2005). "All three Ca2+-binding loops of photoproteins bind calcium ions: the crystal structures of calcium-loaded apo-aequorin and apo-obelin". Protein Sci. 14 (3): 663–75. doi:10.1110/ps.041142905. PMC 2279293. PMID 15689515.
- ↑ Shimomura O, Inouye S (1996). "Titration of recombinant aequorin with calcium chloride". Biochem. Biophys. Res. Commun. 221 (1): 77–81. doi:10.1006/bbrc.1996.0548. PMID 8660347.
- ↑ Ohmiya Y, Kurono S, Ohashi M, Fagan TF, Tsuji FI (1993). "Mass spectrometric evidence for a disulfide bond in aequorin regeneration". FEBS Lett. 332 (3): 226–8. doi:10.1016/0014-5793(93)80637-a. PMID 8405461.
- ↑ Inouye S (2004). "Blue fluorescent protein from the calcium-sensitive photoprotein aequorin is a heat-resistant enzyme, catalyzing the oxidation of coelenterazine". FEBS Lett. 577 (1-2): 105–10. doi:10.1016/j.febslet.2004.09.078. PMID 15527769.
- ↑ Prasher D, McCann RO, Cormier MJ (1985). "Cloning and expression of the cDNA coding for aequorin, a bioluminescent calcium-binding protein". Biochem. Biophys. Res. Commun. 126 (3): 1259–68. doi:10.1016/0006-291X(85)90321-3. PMID 2579647.
- ↑ Inouye S, Noguchi M, Sakaki Y, Takagi Y, Miyata T, Iwanaga S, Miyata T, Tsuji FI (1985). "Cloning and sequence analysis of cDNA for the luminescent protein aequorin". Proc. Natl. Acad. Sci. U.S.A. 82 (10): 3154–8. doi:10.1073/pnas.82.10.3154. PMC 397733. PMID 3858813.
- ↑ Masuda H, Takenaka Y, Shikamoto Y, Kagawa M, Mizuno H, Tsuji FI (2003). "Chromatography of isoforms of recombinant apoaequorin and method for the preparation of aequorin". Protein Expr. Purif. 31 (2): 181–7. doi:10.1016/s1046-5928(03)00186-4. PMID 14550635.
- ↑ Harvey EN (1926). "Oxygen and Luminescence, with a Description of Methods for Removing Oxygen from Cells and Fluids". Biological Bulletin. 51 (2): 89–97. doi:10.2307/1536540.
- ↑ Harvey, E.N. "Bioluminescence" Academic Press., 1952.
- ↑ Shimomura O, Johnson FH (1975). "Regeneration of the photoprotein aequorin". Nature. 256 (5514): 236–238. doi:10.1038/256236a0. PMID 239351.
- ↑ Shimomura O, Johnson FH, Morise H (1974). "Mechanism of the luminescent intramolecular reaction of aequorin". Biochemistry. 13 (16): 3278–3286. doi:10.1021/bi00713a016. PMID 4152180.
- ↑ Shimomura O, Johnson FH (1970). "Calcium binding, quantum yield, and emitting molecule in aequorin bioluminescence". Nature. 227 (5265): 1356–7. doi:10.1038/2271356a0. PMID 4393938.
- ↑ Inouye S, Sasaki S (2006). "Blue fluorescent protein from the calcium-sensitive photoprotein aequorin: catalytic properties for the oxidation of coelenterazine as an oxygenase". FEBS Lett. 580 (8): 1977–82. doi:10.1016/j.febslet.2006.02.065. PMID 16545379.
- ↑ Shimomura O, Johnson FH (1975). "Chemical nature of bioluminescence systems in coelenterates". Proceedings of the National Academy of Sciences. 72 (4): 1546–1549. doi:10.1073/pnas.72.4.1546. PMC 432574. PMID 236561.
- ↑ Shimomura O, Inouye S, Musicki B, Kishi Y (1990). "Recombinant aequorin and recombinant semi-synthetic aequorins. Cellular Ca2+ ion indicators". Biochem. J. 270 (2): 309–12. PMC 1131721. PMID 2400391.
- ↑ Ridgway EB, Ashley CC (1967). "Calcium transients in single muscle fibers". Biochem. Biophys. Res. Commun. 29 (2): 229–34. doi:10.1016/0006-291x(67)90592-x. PMID 4383681.
- ↑ Cheung CY, Webb SE, Meng A, Miller AL (2006). "Transient expression of apoaequorin in zebrafish embryos: extending the ability to image calcium transients during later stages of development". Int. J. Dev. Biol. 50 (6): 561–9. doi:10.1387/ijdb.062151cc. PMID 16741871.
- ↑ Rembold CM, Kendall JM, Campbell AK (January 1997). "Measurement of changes in sarcoplasmic reticulum [Ca2+] in rat tail artery with targeted apoaequorin delivered by an adenoviral vector". Cell Calcium. 21 (1): 69–79. doi:10.1016/s0143-4160(97)90098-1. PMID 9056079.
- ↑ Yamano K, Mori K, Nakano R, Kusunoki M, Inoue M, Satoh M (2007). "Identification of the functional expression of adenosine A3 receptor in pancreas using transgenic mice expressing jellyfish apoaequorin". Transgenic Res. 16 (4): 429–35. doi:10.1007/s11248-007-9084-0. PMID 17387626.
- ↑ Sheu YA, Kricka LJ, Pritchett DB (1993). "Measurement of intracellular calcium using bioluminescent aequorin expressed in human cells". Anal. Biochem. 209 (2): 343–7. doi:10.1006/abio.1993.1132. PMID 8470808.
- ↑ Mithöfer A, Mazars C (2002). "Aequorin-based measurements of intracellular Ca2+ signatures in plant cells". Biol. Proced. Online. 4: 105–18. doi:10.1251/bpo40. PMC 145563. PMID 12734562.
- ↑ Blinks JR, Wier WG, Hess P, Prendergast FG (1982). "Measurement of Ca2+ concentrations in living cells". Prog Biophys Mol Biol. 40 (1–2): 1–114. doi:10.1016/0079-6107(82)90011-6. PMID 6758036.
- ↑ Montero M, Brini M, Marsault R, Alvarez J, Sitia R, Pozzan T, Rizzuto R (1995). "Monitoring dynamic changes in free Ca2+ concentration in the endoplasmic reticulum of intact cells". EMBO J. 14 (22): 5467–75. PMC 394660. PMID 8521803.
- ↑ Kendall JM, Badminton MN, Sala-Newby GB, Campbell AK, Rembold CM (1996). "Recombinant apoaequorin acting as a pseudo-luciferase reports micromolar changes in the endoplasmic reticulum free Ca2+ of intact cells". Biochem J. 318: 383–7. PMC 1217633. PMID 8809023.
- ↑ Miyawaki A, Llopis J, Heim R, McCaffery JM, Adams JA, Ikura M, Tsien RY (1997). "Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin". Nature. 388 (6645): 882–7. doi:10.1038/42264. PMID 9278050.
- ↑ Heim N, Griesbeck O (2004). "Genetically encoded indicators of cellular calcium dynamics based on troponin C and green fluorescent protein". J Biol Chem. 279 (14): 14280–6. doi:10.1074/jbc.M312751200. PMID 14742421.