Xylose isomerase
| xylose isomerase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
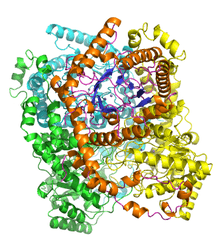 D-Xylose isomerase tetramer from Streptomyces rubiginosus PDB 2glk.[1] One monomer is coloured by secondary structure to highlight the TIM barrel architecture. | |||||||||
| Identifiers | |||||||||
| EC number | 5.3.1.5 | ||||||||
| CAS number | 9023-82-9 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / EGO | ||||||||
| |||||||||
In enzymology, a xylose isomerase (EC 5.3.1.5) is an enzyme that catalyzes the interconversion of D-xylose and D-xylulose. This enzyme belongs to the family of isomerases, specifically those intramolecular oxidoreductases interconverting aldoses and ketoses. The isomerase has now been observed in nearly a hundred species of bacteria. Xylose-isomerases are also commonly called fructose-isomerases due to their ability to convert glucose to fructose. The systematic name of this enzyme class is D-xylose aldose-ketose-isomerase. Other names in common use include D-xylose isomerase, D-xylose ketoisomerase, and D-xylose ketol-isomerase.
History
The activity of D-xylose isomerase was first observed by Mitsuhashi and Lampen in 1953 in the bacterium Lactobacillus pentosus.[2] Artificial production through transformed E.coli have also been successful.[3] D-xylose isomerase was first crystallized from the Lactobacillus brevis strain.[4] In 1957, the D-xylose isomerase activity on D-glucose conversion to D-fructose was noted by Kooi and Marshall.[5] It is now known that isomerases have broad substrate specificity. Most pentoses and some hexoses are all substrates for D-xylose isomerase. Some examples include: D-ribose, L-arabinose, L-rhanmose, and D-allose.[6]
Conversion of glucose to fructose by xylose isomerase was first patented in the 1960s, however, the process was not industrially viable as the enzymes were suspended in solution, and recycling the enzyme was problematic.[6] An immobile xylose isomerase that was fixed on a solid surface was first developed in Japan by Takanashi.[4][6] Due to the development of this technology and its application to produce enriched fructose corn syrup(EFCS), sucrose in United States' soft drinks have been almost completely been substituted by EFCS's.[4]
Function
This enzyme participates in pentose and glucuronate interconversions and fructose and mannose metabolism. The most bio-available sugars according to the International Society of Rare Sugars are: glucose, galactose, mannose, fructose, xylose, ribose, and L-arabinose. Twenty hexoses and nine pentoses, including xylulose, were considered to be "rare sugars". Hence D-xylose isomerase is used to produce these rare sugars which have very important applications in biology despite their low abundance.[7]
Characterization
Xylose isomerase that can be isolated from red Chinese rice wine, which contains the bacterium Lactobacillus xylosus.[8] This bacterium was mistakenly classified as a L. plantarum, which normally grows on the sugar L-arabinose, and rarely grown on D-xylose. L. xylosus was recognized to be distinct for its ability to grow on D-xylose.[9] Xylose isomerase in L. xylosus has a molecular weight of about 183000 Daltons.[10] Its optimum growth pH is about 7.5 for the L. lactis, however strains such as the L.brevis xylose enzyme prefer a more alkaline environment. The L. lactis strain is stable over the pH range of 6.5 to 11.0, and the L. brevis enzyme, which is less tolerant of pH changes, show activity over the pH range of 5.7-7.0.[10][10] Thermal tests were also done by Kei Y. and Noritaka T. and the xylose isomerase was found to be thermally stable to about 60 degrees Celsius[10]
Xylose isomerase active site and mechanism
Xylose isomerase has a structure that is based on eight alpha/beta barrels that create an active site holding two divalent magnesium ions. Xylose isomerase enzymes exhibit a TIM barrel fold with the active site in the centre of the barrel and a tetrameric quaternary structure.[11] PDB structures are available in the links in the infobox to the right. The protein is a tetramer where paired barrels are nearly coaxial, which form two cavities in which the divalent metals are both bound to one of the two cavities. The metals are in an octahedral geometry. Metal site 1 binds the substrate tightly, while metal site two binds the substrate loosely. Both share an acid residue Glutamic acid 216 of the enzyme that bridges the two cations. Two basic amino acids surround the negative charged ligands to neutralize them. The second cavity faces the metal cavity and both cavities share the same access route. The second cavity is hydrophobic in nature, and has an important histidine residue that is activated by an aspartate residue that is hydrogen bonded to it. This histidine residue is important in the isomerization of glucose.[12]
In the isomerization of glucose, Histidine 53 is used to catalyze the proton transfer of O1 to O5; the diagram for the ring opening mechanism is shown below. The first metal, mentioned earlier, coordinates to O3 and O4, and is used to dock the substrate.[12]

In the isomerization of xylose, crystal data has shown that xylose sugar binds to the enzyme in an open chain conformation. Metal 1 binds to O2 and O4, and once bound, metal 2 binds to O1 and O2 in the transition state, and these interactions along with a lysine residue help catalyze the hydride shift necessary for isomerization. The transition state consists of a high energy carbonium ion that is stabilized through all the metal interactions with the sugar substrate.[12]

Application in industry
The most widely used application of this enzyme is in the conversion of glucose to fructose to produce high fructose corn syrup (HFCS).[4][6] There are three general steps in producing HFCS from starch,
- enzymatic degradation of the starch using α-amylase. Also known as liquification.[4]
- further degradation using glucoamylase and a debranching enzyme.[4]
- Production of fructose through xylose isomerase[4]
The process is done in elevated temperatures, where the xylose-isomerase enzyme is adsorbed onto polyacrylamide gels (carrier material), and the enzymes produce batches and are reused. In a report done in 1998, 80% of the enzyme reactivity was retained after 40 days of catalysis at 60 °C.[4] It has been shown that immobilized xylose-isomerases can be used in processes with elevated temperatures with a half life of ten-thousand years.[13] This fact has opened up hope for the enzyme catalyzing the production of rare sugar. The carrier material holds a low percentage of enzyme, and a problem in making the production of rare sugars through non-conventional substrates is that the affinity for these substrates are 2-500 times slower. The production of these rare sugars is not feasible today, as it requires large volumes of carrier material with the enzyme. Cross-linking xylose-isomerase enzymes gives a higher density on the carrier. This new technology has opened up hopes for the reduction of prices of rare sugars through xylose-isomerase.[6] Xylose-isomerases can also be used in the production of biofuels. After their heterologous expression, xylulose can be produced from xylose and fermented into ethanol using yeasts.[4][14]
References
- ↑ Katz, A. K.; Li, X.; Carrell, H. L.; Hanson, B. L.; Langan, P.; Coates, L.; Schoenborn, B. P.; Glusker, J. P.; Bunick, G. J. (2006). "Locating active-site hydrogen atoms in D-xylose isomerase: Time-of-flight neutron diffraction". Proceedings of the National Academy of Sciences. 103 (22): 8342–8347. doi:10.1073/pnas.0602598103. PMC 1482496
 . PMID 16707576.
. PMID 16707576. - ↑ Mitsuhashi, S.; Lampen, J. (1953). "Conversion of D-xylose to D-xylulose in extracts of Lactobacillus pentosus". Biological Chemistry. 204: 1011.
- ↑ Schomburg, Dietmar (2001). Handbook of Enzymes. New York: Springer. pp. 259–260. Retrieved 12 March 2015.
- 1 2 3 4 5 6 7 8 9 Pastinen, Ossi. "Xylose Isomerase" (PDF). NOPPA. Retrieved 13 March 2015.
- ↑ Marshall, Richard; Kooi, Earl (1957). "Enzymic conversion of D-glucose to D-fructose". Science. 125: 648–9. doi:10.1126/science.125.3249.648. PMID 13421660.
- 1 2 3 4 5 Jokela, Jouni; Pastinen, Ossi (2002). "Isomerization of pentose and hexose sugars by an enzyme reactor packed with cross-linked xylose isomerase crytals". Enzyme and Microbial Technology. 31: 67–76. doi:10.1016/s0141-0229(02)00074-1.
- ↑ Beerens, K. (2012). "Enzymes for the biocatalytic production of rare sugars". J. Ind. Microbiol. Biotechnol. 39: 823–834. doi:10.1007/s10295-012-1089-x.
- ↑ Kitahara, K. (1966). "Studies on Lactic Acid Bacteria". Nyusankin no Kenkyu: 67~69.
- ↑ Buchanan, R.E.; Gibbons, N.E. (1974). Bergey's Manual of Determining Bacteriology (8 ed.). Baltimore: The Williams and Wilkins Co. p. 584.
- 1 2 3 4 Yamanaka, Kei; Takahara, Noritaka (1977). "Purification and Properties of D-Xylose Isomerase from Lactobacillus xylosus". Agric. Biol. Chem. 41: 1909–1915. doi:10.1271/bbb1961.41.1909.
- ↑ Deprecated services < EMBL-EBI
- 1 2 3 Blow, David; Collyer, Charles; Goldberg, Jonathan; Smart, Oliver (1992). "Structure and Mechanism of D-xylose Isomerase". Faraday Discussions. 93: 67. doi:10.1039/fd9929300067.
- ↑ Volkin, D; Klibanow, A. (1988). "Mechanism of thermoinacctivation of immobilized glucose isomerase". Biotechnol Bioeng. 33: 1104–1111. doi:10.1002/bit.260330905.
- ↑ Maris, Van; Antonius, et.al (2007). "Development of Efficient Xylose Fermentation in Saccharomyces Cerevisiae: Xylose Isomerase as a key component". Adv. Biochem. Engin/Biotechnol. 108: 179.
Further reading
- Hochster RM, Watson RW (1954). "Enzymatic isomerization of D-xylose to D-xylulose". Arch. Biochem. Biophys. 48 (1): 120–9. doi:10.1016/0003-9861(54)90313-6. PMID 13125579.
- Slein MW (1955). "Xylose isomerase from Pasteurella pestis, strain A-1122". J. Am. Chem. Soc. 77 (6): 1663–1667. doi:10.1021/ja01611a074.
- Yamanaka K (1968). "Purification, crystallization and properties of the D-xylose isomerase from Lactobacillus brevis". Biochim. Biophys. Acta. 151 (3): 670–80. doi:10.1016/0005-2744(68)90015-6. PMID 5646045.