Haplogroup C-M217
| Haplogroup C-M217 C2 (previously C3) [1] | |
|---|---|
 | |
| Possible time of origin | 11,900 ± 4,800 years before present[2] |
| Possible place of origin | Probably Central Asia or East Asia |
| Ancestor | C M130 |
| Descendants | C-M93 (C2a); C-CTS117 (C2b); C-P53.1 (C2c); C-P62 (C2d); C-F2613/Z1338 (C2e) |
| Defining mutations | M217, P44, PK2 |
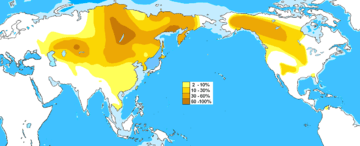
| Highest frequencies | Oroqen 61%[3]-91%,[4] Evens 5%[5]-74%,[6] Evenks 44%[4]-71%,[2][5] Buryats 7%[7]-84%,[8] Mongolians 52%[4]-54%,[3] Kazakhs 40%[4]-60.7%,[9] Tanana 42%,[10] Hazaras 40%,[11] Nivkhs 38%,[8] Koryaks 33%,[2][5] Daur 31%,[3] Yukaghir 31%,[12] Sibe 27%,[3] Manchu 26%[3]-27%,[4] Altai 22%[6]-24%,[4] Hezhe 22%,[3] Uzbeks 20%,[4] Tujia 18%,[4] Hani 18%,[3] Cheyenne 16%,[10] Apache 15%,[10] Tuvans 15%,[12] Ainu 12.5%[8]-25%,[6] Koreans 13%[4]-16%, 20% [3] Hui 11%,[3][4] Sioux 11%,[10] Nogais 14%,[13] Crimean Tatars 9%,[13] Han Chinese 0%-23.5%,[14][15] Vietnamese 4.3%-12.5%[15] |
Haplogroup C-M217, also known as C2 (and previously as C3),[1] is a Y-chromosome DNA haplogroup. It is the most frequently occurring branch of the wider Haplogroup C (M130).
M217 is found at high frequencies among Central Asian peoples, indigenous Siberians, and some Native peoples of North America. In particular, males belonging to peoples such as the Buryats, Evens, Evenks, Kazakhs, Mongolians and Udegeys have high levels of M217.[4][6][16]
One particular haplotype within Haplogroup C2 (M217) has received a great deal of attention, because of the possibility that it may represent direct patrilineal descent from Genghis Khan,[17] one of his male-line ancestors, and/or other males belonging to the Borjigin clan. However, it is not certain that the Y-DNA haplogroup possessed by Genghis Khan was C2 (M217). Haplogroup C3c has been identified as a possible marker of the Manchu Aisin Gioro and is found in ten different ethnic minorities in northern China, but completely absent from Han Chinese.[18][19]
Origin
Haplogroup C-M217 is believed to have originated approximately 7,100 to 16,700 years before present[2] in eastern or central Asia. Its closest phylogenetic relatives are found in the general vicinity of South Asia, East Asia, or Oceania.
The extremely broad distribution of Haplogroup C-M217 Y-chromosomes, coupled with the fact that the ancestral paragroup C is not found among any of the modern Siberian or North American populations among whom Haplogroup C-M217 predominates, makes the determination of the geographical origin of the defining M217 mutation exceedingly difficult. The presence of Haplogroup C-M217 at a low frequency but relatively high diversity throughout East Asia and parts of Southeast Asia makes that region one likely source. In addition, the C-M217 haplotypes found with high frequency among North Asian populations appear to belong to a different genealogical branch from the C-M217 haplotypes found with low frequency among East and Southeast Asians, which suggests that the marginal presence of C-M217 among modern East and Southeast Asian populations may not be due to recent admixture from Northeast or Central Asia.[20]
Distribution
Haplogroup C-M217 is the modal haplogroup among Mongolians and most indigenous populations of the Russian Far East, such as the Northern Tungusic peoples, Koryaks, Itelmens, and Nivkhs. The subclade C-P39 is common among males of the indigenous North American peoples whose languages belong to the Na-Dené phylum. The frequency of Haplogroup C-M217 tends to be negatively correlated with distance from Mongolia and the Russian Far East, but it still comprises more than ten percent of the total Y-chromosome diversity among the Manchus, Koreans, Ainu, and some Turkic peoples of Central Asia although in a genetic study in 2004, haplogroup C-M217 was more frequent among Koreans than previously thought (16.5%). It is found at lower frequencies among Turkic peoples of Central Asia except Kazakhs. On the other hand, Mongols and Tungusic peoples have the highest frequencies. It is possible that this haplogroup might have spread into Turks during the Mongol invasions of 13th century as well as previous tribal leagues in the history.[21] Beyond this range of high-to-moderate frequency, which contains mainly the northeast quadrant of Eurasia and the northwest quadrant of North America, Haplogroup C-M217 continues to be found at low frequencies, and it has even been found as far afield as Northwest Europe, Turkey, Pakistan, Nepal[22] and adjacent regions of India,[23][24][25] Vietnam, Maritime Southeast Asia, and the Wayuu people of South America.
In Japan, haplogroup C-M217 has been found relatively frequently among Ainus (2/16=12.5%[8] or 1/4=25%[6]) and among Japanese of the Kyūshū region (4/53=7.5%[6] or 8/104=7.7%[8]). In other regional Japanese populations, the frequency of haplogroup C-M217 hovers around 2%.
The frequency of Haplogroup C-M217 in samples of Han Chinese from various areas has ranged from 0% (0/27 Han from Guangxi) to 23.5% (8/34 Han from Xi'an[15]), with the frequency of this haplogroup in several studies' pools of all Han Chinese samples ranging between 6.0% and 12.0%.[3][4][6][8][14][15] C-M217 also has been found in samples of minority populations from central and southern China, such as Tujia from Jishou, Hunan (9/49 = 18.4%), Hani (6/34 = 17.6%), Miao (2/58 = 3.4%), She (1/34 = 2.9%), Yao (1/35 = 2.9% from Liannan, Guangdong), Tibetans (4/156 = 2.6%), and Taiwanese aborigines (1/48 = 2.1%). (Karafet 2010)(Xue 2006)(Gayden 2007)
In Vietnam, Haplogroup C-M217 has been found in 12.5% (6/48) of a sample of Vietnamese from Hanoi, Vietnam,[15] 11.8% (9/76) of another sample of Kinh ("ethnic Vietnamese") from Hanoi, Vietnam, 8.5% (5/59) of a sample of Cham people from Binh Thuan, Vietnam, 8.3% (2/24) of another sample of Vietnamese from Hanoi,[26] and in 4.3% (3/70) of a sample of Vietnamese from an unspecified location in Vietnam.(Karafet 2010)(He 2012) Haplogroup C-M217 has been found less frequently in other parts of Southeast Asia and nearby areas, including Laos (1/25 = 4.0% Lao from Luang Prabang), Java (1/37 = 2.7%), Nepal (2/77 = 2.6% general population of Kathmandu), Thailand (1/40 = 2.5% Thai, mostly sampled in Chiang Mai), the Philippines (1/48 = 2.1%, 1/64 = 1.6%), and Bali (1/641 = 0.2%). (Gayden 2007)(Karafet 2010)(He 2012)[15]
Although C-M217 is generally found with only low frequency (<5%) in Tibet and Nepal, there may be an island of relatively high frequency of this haplogroup in Meghalaya, India. The indigenous tribes of this state of Northeast India, where they comprise the majority of the local population, speak Khasian languages or Tibeto-Burman languages. A study published in 2007 found C-M217(xM93, P39, M86) Y-DNA in 8.5% (6/71) of a sample of Garos, who primarily inhabit the Garo Hills in the western half of Meghalaya, and in 7.6% (27/353) of a pool of samples of eight Khasian tribes from the eastern half of Meghalaya (6/18 = 33.3% Nongtrai from the West Khasi Hills, 10/60 = 16.7% Lyngngam from the West Khasi Hills, 2/29 = 6.9% War-Khasi from the East Khasi Hills, 3/44 = 6.8% Pnar from the Jaintia Hills, 1/19 = 5.3% War-Jaintia from the Jaintia Hills, 3/87 = 3.4% Khynriam from the East Khasi Hills, 2/64 = 3.1% Maram from the West Khasi Hills, and 0/32 Bhoi from Ri-Bhoi District).(Reddy 2007)
Subclade distribution
The subclades of Haplogroup C-M217 with their defining mutation(s), according to the 2008 ISOGG tree:
C2-M217 Typical of Mongolians, Kazakhs, Buryats, Daurs, Kalmyks, Hazaras, Manchus, Sibes, Oroqens, Koryaks, and Itelmens; with a moderate distribution among other Tungusic peoples, Koreans, Ainus, Han Chinese, Vietnamese, Nivkhs, Altaians, Tuvinians, Uzbeks, Nogais, and Crimean Tatars[3][6][7][8][11][13][16][27]
C2-M93 Observed sporadically in Japanese[11][28]
C2-P39 Found in several indigenous peoples of North America, including some Na-Dené-, Algonquian-, or Siouan-speaking populations[10]
C2-M48 Found frequently in Koryaks and sporadically among Evenks, Evens, and Yukaghirs
C2-M77 Typical of Northern Tungusic peoples, Kazakhs, Oirats, Kalmyks, Outer Mongolians, Yukaghirs, Nivkhs, Itelmens, and Udegeys, with a moderate distribution among other Southern Tungusic peoples, Inner Mongolians, Buryats, Tuvinians, Yakuts, Chukchi, Kyrgyz, Uyghurs, Uzbeks, Karakalpaks, and Tajiks[7][12][29]
C2-M407 Found with low frequency in Bai, Cambodian, Han Chinese, Japanese, Korean,[30] Manchu, Tujia, Uyghur, and Yakut populations[11][14]
C2-P53.1 Found in about 10% of Xinjiang Sibe and with low frequency in Inner Mongolian Mongol and Evenk, Ningxia Hui, Xizang Tibetan, Xinjiang Uyghur, and Gansu Han[14]
C2-P62
Phylogenetics
Phylogenetic history
Prior to 2002, there were in academic literature at least seven naming systems for the Y-Chromosome Phylogenetic tree. This led to considerable confusion. In 2002, the major research groups came together and formed the Y-Chromosome Consortium (YCC). They published a joint paper that created a single new tree that all agreed to use. Later, a group of citizen scientists with an interest in population genetics and genetic genealogy formed a working group to create an amateur tree aiming at being above all timely. The table below brings together all of these works at the point of the landmark 2002 YCC Tree. This allows a researcher reviewing older published literature to quickly move between nomenclatures.
| YCC 2002/2008 (Shorthand) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | (η) | YCC 2002 (Longhand) | YCC 2005 (Longhand) | YCC 2008 (Longhand) | YCC 2010r (Longhand) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C-M216 | 10 | V | 1F | 16 | Eu6 | H1 | C | C* | C | C | C | C | C | C | C | C | C | C |
| C-M8 | 10 | V | 1F | 19 | Eu6 | H1 | C | C1 | C1 | C1 | C1 | C1 | C1 | C1 | C1 | C1 | C1 | C1 |
| C-M38 | 10 | V | 1F | 16 | Eu6 | H1 | C | C2* | C2 | C2 | C2 | C2 | C2 | C2 | C2 | C2 | C2 | C2 |
| C-P33 | 10 | V | 1F | 18 | Eu6 | H1 | C | C2a | C2a | C2a1 | C2a1 | C2a | C2a | C2a1 | C2a1 | C2a1 | removed | removed |
| C-P44 | 10 | V | 1F | 17 | Eu6 | H1 | C | C3* | C3 | C3 | C3 | C3 | C3 | C3 | C3 | C3 | C3 | C3 |
| C-M93 | 10 | V | 1F | 17 | Eu6 | H1 | C | C3a | C3a | C3a | C3a | C3a | C3a | C3a | C3a | C3a | C3a | C3a1 |
| C-M208 | 10 | V | 1F | 17 | Eu6 | H1 | C | C3b | C2b | C2a | C2a | C2b | C2b | C2a | C2a | C2a | C2a | C2a |
| C-M210 | 36 | V | 1F | 17 | Eu6 | H1 | C | C3c | C2c | C4a | C4a | C4b | C4b | C4a | C4a | C4a | C4a | C4a |
Research publications
The following research teams per their publications were represented in the creation of the YCC tree.
See also
Genetics
- African admixture in Europe
- Genetic genealogy
- Haplogroup
- Haplotype
- Human Y-chromosome DNA haplogroup
- Molecular phylogeny
- Paragroup
- Subclade
- Y-chromosome haplogroups by populations
- Y-DNA haplogroups by ethnic groups
- Y-DNA haplogroups by populations of East and Southeast Asia
- Y-DNA haplogroups in Oceanian populations
Y-DNA C subclades
- Mega-Haplogroup CF
- Mega-Haplogroup CT
- C-M130
- C-M208
- C-M210
- C-M216
- C-M217
- C-M38
- C-M8
- C-M93
- C-P33
- C-P44
Y-DNA backbone tree
| Phylogenetic tree of human Y-chromosome DNA haplogroups [χ 1][χ 2] | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-chromosomal Adam" | |||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | ||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | ||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | ||||||||||||||||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | H | ||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | LT [χ 5] | K2 | ||||||||||||||||||||||||||||||||||||||||||||||
| L | T [χ 6] | NO [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | |||||||||||||||||||||||||||||||||||||||||||
| N | O | K2b1 [χ 10] | P | ||||||||||||||||||||||||||||||||||||||||||||||
| K2b1a [χ 11] | K2b1b | K2b1c | M | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||
| K2b1a1 | K2b1a2 | K2b1a3 | S [χ 12] | Q | R | ||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
References
- 1 2 ISOGG, 2015 "Y-DNA Haplogroup C and its Subclades – 2015" (15 September 2015).
- 1 2 3 4 Karafet TM, Osipova LP, Gubina MA, Posukh OL, Zegura SL, Hammer MF (December 2002). "High levels of Y-chromosome differentiation among native Siberian populations and the genetic signature of a boreal hunter-gatherer way of life". Hum. Biol. 74 (6): 761–89. doi:10.1353/hub.2003.0006. PMID 12617488.
- 1 2 3 4 5 6 7 8 9 10 11 Xue Y, Zerjal T, Bao W, et al. (April 2006). "Male demography in East Asia: a north-south contrast in human population expansion times". Genetics. 172 (4): 2431–9. doi:10.1534/genetics.105.054270. PMC 1456369
 . PMID 16489223.
. PMID 16489223. - 1 2 3 4 5 6 7 8 9 10 11 12 Karafet T, Xu L, Du R, et al. (September 2001). "Paternal population history of East Asia: sources, patterns, and microevolutionary processes". Am. J. Hum. Genet. 69 (3): 615–28. doi:10.1086/323299. PMC 1235490. PMID 11481588.
- 1 2 3 Pakendorf B, Novgorodov IN, Osakovskij VL, Stoneking M (July 2007). "Mating patterns amongst Siberian reindeer herders: inferences from mtDNA and Y-chromosomal analyses". Am. J. Phys. Anthropol. 133 (3): 1013–27. doi:10.1002/ajpa.20590. PMID 17492671.
- 1 2 3 4 5 6 7 8 Hammer MF, Karafet TM, Park H, et al. (2006). "Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes". J. Hum. Genet. 51 (1): 47–58. doi:10.1007/s10038-005-0322-0. PMID 16328082.
- 1 2 3 Lell JT, Sukernik RI, Starikovskaya YB, et al. (January 2002). "The dual origin and Siberian affinities of Native American Y chromosomes". Am. J. Hum. Genet. 70 (1): 192–206. doi:10.1086/338457. PMC 384887. PMID 11731934.
- 1 2 3 4 5 6 7 Tajima, Atsushi; Hayami, Masanori; Tokunaga, Katsushi; Juji, T; Matsuo, M; Marzuki, S; Omoto, K; Horai, S (2004). "Genetic origins of the Ainu inferred from combined DNA analyses of maternal and paternal lineages". Journal of Human Genetics. 49 (4): 187–193. doi:10.1007/s10038-004-0131-x. PMID 14997363.
- ↑ Dulik MC, Osipova LP, Schurr TG (2011). "Y-chromosome variation in Altaian Kazakhs reveals a common paternal gene pool for Kazakhs and the influence of Mongolian expansions". PLoS ONE. 6 (3): e17548. doi:10.1371/journal.pone.0017548. PMC 3055870. PMID 21412412.
- 1 2 3 4 5 Zegura SL, Karafet TM, Zhivotovsky LA, Hammer MF (January 2004). "High-resolution SNPs and microsatellite haplotypes point to a single, recent entry of Native American Y chromosomes into the Americas". Mol. Biol. Evol. 21 (1): 164–75. doi:10.1093/molbev/msh009. PMID 14595095.
- 1 2 3 4 Sengupta S, Zhivotovsky LA, King R, et al. (February 2006). "Polarity and temporality of high-resolution y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic influence of Central Asian pastoralists". Am. J. Hum. Genet. 78 (2): 202–21. doi:10.1086/499411. PMC 1380230. PMID 16400607.
- 1 2 3 Pakendorf B, Novgorodov IN, Osakovskij VL, Danilova AP, Protod'jakonov AP, Stoneking M (October 2006). "Investigating the effects of prehistoric migrations in Siberia: genetic variation and the origins of Yakuts". Hum. Genet. 120 (3): 334–53. doi:10.1007/s00439-006-0213-2. PMID 16845541.
- 1 2 3 Marchani EE, Watkins WS, Bulayeva K, Harpending HC, Jorde LB (2008). "Culture creates genetic structure in the Caucasus: autosomal, mitochondrial, and Y-chromosomal variation in Daghestan". BMC Genet. 9: 47. doi:10.1186/1471-2156-9-47. PMC 2488347. PMID 18637195.
- 1 2 3 4 Zhong, Hua; Shi, Hong; Xue-, XB; Qi, Bin; Jin, L; Ma, RZ; Su, B (2010). "Global distribution of Y-chromosome haplogroup C reveals the prehistoric migration routes of African exodus and early settlement in East Asia". Journal of Human Genetics. 55 (7): 428–35. doi:10.1038/jhg.2010.40. PMID 20448651.
- 1 2 3 4 5 6 Kim SH, Kim KC, Shin DJ, et al. (2011). "High frequencies of Y-chromosome haplogroup O2b-SRY465 lineages in Korea: a genetic perspective on the peopling of Korea". Investig Genet. 2 (1): 10. doi:10.1186/2041-2223-2-10. PMC 3087676. PMID 21463511.
- 1 2 Wells RS, Yuldasheva N, Ruzibakiev R, et al. (August 2001). "The Eurasian heartland: a continental perspective on Y-chromosome diversity". Proc. Natl. Acad. Sci. U.S.A. 98 (18): 10244–9. doi:10.1073/pnas.171305098. PMC 56946. PMID 11526236.
- ↑ Zerjal T, Xue Y, Bertorelle G, et al. (March 2003). "The genetic legacy of the Mongols". Am. J. Hum. Genet. 72 (3): 717–21. doi:10.1086/367774. PMC 1180246. PMID 12592608. as PDF
- ↑ Xue, Y; Zerjal, T; Bao, W; Zhu, S; Lim, SK; Shu, Q; Xu, J; Du, R; Fu, S; Li, P; Yang, H; Tyler-Smith, C (2015-09-28). "Recent Spread of a Y-Chromosomal Lineage in Northern China and Mongolia". Am. J. Hum. Genet. 77: 1112–6. doi:10.1086/498583. PMC 1285168. PMID 16380921.
- ↑ Xue, Y; Zerjal, T; Bao, W; Zhu, S; Lim, SK; Shu, Q; Xu, J; Du, R; Fu, S; Li, P; Yang, H; Tyler-Smith, C (2005). "Recent Spread of a Y-Chromosomal Lineage in Northern China and Mongolia". The American Journal of Human Genetics. 77: 1112–1116. doi:10.1086/498583. PMC 1285168. PMID 16380921. Retrieved 2015-11-26.
- ↑ Redd AJ, Roberts-Thomson J, Karafet T, et al. (April 2002). "Gene flow from the Indian subcontinent to Australia: evidence from the Y chromosome". Curr. Biol. 12 (8): 673–7. doi:10.1016/S0960-9822(02)00789-3. PMID 11967156. as PDF
- ↑ KZ DNA Project, FTDNA
- ↑ Gayden T, Cadenas AM, Regueiro M, et al. (May 2007). "The Himalayas as a directional barrier to gene flow". Am. J. Hum. Genet. 80 (5): 884–94. doi:10.1086/516757. PMC 1852741. PMID 17436243. 2/77=2.6% C-M217 in a sample of the general population of Kathmandu.
- ↑ Fornarino S, Pala M, Battaglia V, et al. (2009). "Mitochondrial and Y-chromosome diversity of the Tharus (Nepal): a reservoir of genetic variation". BMC Evol. Biol. 9: 154. doi:10.1186/1471-2148-9-154. PMC 2720951. PMID 19573232. 1/26=3.8% C-M217 in a sample of Hindu Indians from the Terai.
- ↑ Reddy BM, Langstieh BT, Kumar V, et al. (2007). "Austro-Asiatic tribes of Northeast India provide hitherto missing genetic link between South and Southeast Asia". PLoS ONE. 2 (11): e1141. doi:10.1371/journal.pone.0001141. PMC 2065843. PMID 17989774. Haplogroup C-M217 in 8.5% of a sample of 71 Garos and 7.7% of a pool of eight samples of Khasians totalling 353 individuals
- ↑ Kivisild T, Rootsi S, Metspalu M, et al. (February 2003). "The genetic heritage of the earliest settlers persists both in Indian tribal and caste populations". Am. J. Hum. Genet. 72 (2): 313–32. doi:10.1086/346068. PMC 379225. PMID 12536373. C-M217 in 1/31=3.2% of a sample from West Bengal.
- ↑ Jean A Trejaut, Estella S Poloni, Ju-Chen Yen, et al., "Taiwan Y-chromosomal DNA variation and its relationship with Island Southeast Asia." BMC Genetics 2014, 15:77 http://www.biomedcentral.com/1471-2156/15/77
- ↑ Nasidze I, Quinque D, Dupanloup I, Cordaux R, Kokshunova L, Stoneking M (December 2005). "Genetic evidence for the Mongolian ancestry of Kalmyks". Am. J. Phys. Anthropol. 128 (4): 846–54. doi:10.1002/ajpa.20159. PMID 16028228.
- ↑ Underhill PA, Shen P, Lin AA, et al. (November 2000). "Y chromosome sequence variation and the history of human populations". Nat. Genet. 26 (3): 358–61. doi:10.1038/81685. PMID 11062480.
- ↑ Khar'kov VN, Stepanov VA, Medvedev OF, et al. (2008). "[The origin of Yakuts: analysis of Y-chromosome haplotypes]". Mol. Biol. (Mosk.) (in Russian). 42 (2): 226–37. PMID 18610830.
- ↑ Park, Jin; Lee, Young; Kim, Young; -1#Myung, Hwan Na; et al. (2013). "Y-SNP miniplexes for East Asian Y-chromosomal haplogroup determination in degraded DNA". Forensic Science International: Genetics. 7: 75–81. doi:10.1016/j.fsigen.2012.06.014.