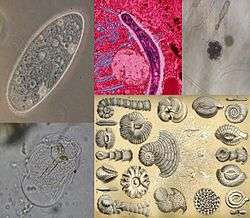
Protist
| Protist Temporal range: Neoproterozoic – Recent | |
|---|---|
 | |
| Scientific classification | |
| Domain: | Eukarya |
classification varies | |
Protist (/ˈproʊtᵻst/) is an informal term for any eukaryotic organism that is not an animal, plant or fungus. The protists do not form a natural group, or clade, but are often grouped together for convenience, like algae or invertebrates. In some systems of biological classification, such as the popular 5-kingdom scheme proposed by Robert Whittaker in 1969, the protists make up a kingdom called Protista, composed of "organisms which are unicellular or unicellular-colonial and which form no tissues."[2][3][upper-alpha 1]
Besides their relatively simple levels of organization, protists do not necessarily have much in common.[6] When used, the term “protists” is now considered to mean similar-appearing but diverse taxa that are not related through an exclusive common ancestor, and have different life cycles, trophic levels, modes of locomotion, and cellular structures.[7][8] In the classification system of Lynn Margulis, the term protist is reserved for microscopic organisms, while the more inclusive term Protoctista is applied to a biological kingdom which includes certain large multicellular eukaryotes, such as kelp, red algae and slime molds.[9] Other workers use the term protist more broadly, to encompass both microbial eukaryotes and macroscopic organisms that do not fit into the other traditional kingdoms.
In cladistic systems, there are no equivalents to the taxa Protista or Protoctista, both terms referring to a paraphyletic group which spans the entire eukaryotic tree of life. In cladistic classification, the contents of Protista are distributed among various supergroups (SAR, Archaeplastida, Excavata, Opisthokonta, etc.) and "Protista", ''Protoctista'' and "Protozoa" are considered obsolete. However, the term "protist" continues to be used informally as a catch-all term for eukaryotic microorganisms. For example, the phrase "protist pathogen" may be used to denote any disease-causing microbe which is not bacteria, virus, viroid or metazoa.[10]
Subdivisions
The term protista was first used by Ernst Haeckel in 1866. Protists were traditionally subdivided into several groups based on similarities to the "higher" kingdoms such as:
Protozoa the unicellular "animal-like" (heterotrophic/parasitic) protozoa which was further sub-divided based on motility such as (flagellated) Flagellata, (ciliated) Ciliophora, (phagocytic) amoeba and spore-forming Sporozoans
Protophyta the "plant-like" (autotrophic) protophyta (mostly unicellular algae)
Molds the "fungus-like" (saprophytic) slime molds and water molds.
Some protists, sometimes called ambiregnal protists, have been considered to be both protozoa and algae or fungi (e.g., slime molds and flagellated algae), and names for these have been published under either or both of the ICN and the ICZN.[11][12] Conflicts, such as these – for example the dual-classification of Euglenids and Dinobryons, which are mixotrophic – is an example of why the kingdom Protista was adopted.
These traditional subdivisions, largely based on superficial commonalities, have been replaced by classifications based on phylogenetics (evolutionary relatedness among organisms). Molecular analyses in modern taxonomy have been used to redistribute former members of this group into diverse and sometimes distantly related phyla. For instance, the water molds are now considered to be closely related to photosynthetic organisms such as Brown algae and Diatoms, the slime molds are grouped mainly under Amoebozoa, and the Amoebozoa itself includes only a subset of "Amoeba" group, and significant number of erstwhile "Amoeboid" genera are distributed among Rhizarians and other Phyla.
However, the older terms are still used as informal names to describe the morphology and ecology of various protists. For example, the term protozoa is used to refer to heterotrophic species of protists that do not form filaments.
Classification
Historical classifications
Among the pioneers in the study of the protists, which were almost ignored by Linnaeus except for some genera (e.g., Vorticella, Chaos, Volvox, Corallina, Conferva, Ulva, Chara, Fucus)[13][14] were Leeuwenhoek, O. F. Müller, C. G. Ehrenberg and Félix Dujardin.[15] The first groups used to classify microscopic organism were the Animalcules and the Infusoria.[16] In 1817, the German naturalist Georg August Goldfuss introduced the word Protozoa to refer to organisms such as ciliates and corals.[17] After the cell theory of Schwann and Schleiden (1838–39), this group was modified in 1848 by Carl von Siebold to include only animal-like unicellular organisms, such as foraminifera and amoebae. The formal taxonomic category Protoctista was first proposed in the early 1860s by John Hogg, who argued that the protists should include what he saw as primitive unicellular forms of both plants and animals. He defined the Protoctista as a "fourth kingdom of nature", in addition to the then-traditional kingdoms of plants, animals and minerals.[17] The kingdom of minerals was later removed from taxonomy in 1866 by Ernst Haeckel, leaving plants, animals, and the protists (Protista), defined as a “kingdom of primitive forms”.[18]
In 1938, Herbert Copeland resurrected Hogg's label, arguing that Haeckel's term Protista included anucleated microbes such as bacteria, which the term "Protoctista" (literally meaning "first established beings") did not. In contrast, Copeland's term included nucleated eukaryotes such as diatoms, green algae and fungi.[19] This classification was the basis for Whittaker's later definition of Fungi, Animalia, Plantae and Protista as the four kingdoms of life.[4] The kingdom Protista was later modified to separate prokaryotes into the separate kingdom of Monera, leaving the protists as a group of eukaryotic microorganisms.[20] These five kingdoms remained the accepted classification until the development of molecular phylogenetics in the late 20th century, when it became apparent that neither protists nor monera were single groups of related organisms (they were not monophyletic groups).[21]
Modern classifications
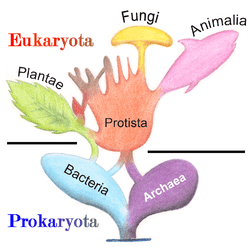
Many systematists today do not treat Protista as a formal taxon, but the term "protist" is still commonly used for convenience in two ways.[22] The most popular contemporary definition is a phylogenetic one, that identifies a paraphyletic group:[23] a protist is any eukaryote that is not an animal, (land) plant, or (true) fungus; this definition[24] excludes many unicellular groups, like the Microsporidia (fungi), many Chytridiomycetes (fungi), and yeasts (fungi), and also a non-unicellular group included in Protista in the past, the Myxozoa (animal).[25] Some systematists judge paraphyletic taxa acceptable, and use Protista in this sense as a formal taxon (as found in some secondary textbooks, for pedagogical purpose).
The other definition describes protists primarily by functional or biological criteria: protists are essentially those eukaryotes that are never multicellular,[22] that either exist as independent cells, or if they occur in colonies, do not show differentiation into tissues (but vegetative cell differentiation may occur restricted to sexual reproduction, alternate vegetative morphology, and quiescent or resistant stages, such as cysts);[26] this definition excludes many brown, many red and some green algae, which may have tissues.
The taxonomy of protists is still changing. Newer classifications attempt to present monophyletic groups based on morphological (especially ultrastructural),[27][28][29] biochemical (chemotaxonomy)[30][31] and DNA sequence (molecular research) information.[32][33] However, there are sometimes discordances between molecular and morphological investigations; these can be categorized as two types: (i) one morphology, multiple lineages (e.g. morphological convergence, cryptic species) and (ii) one lineage, multiple morphologies (e.g. phenotypic plasticity, multiple life-cycle stages).[34]
Because the protists as a whole are paraphyletic, new systems often split up or abandon the kingdom, instead treating the protist groups as separate lines of eukaryotes. The recent scheme by Adl et al. (2005)[26] does not recognize formal ranks (phylum, class, etc.) and instead treats groups as clades of phylogenetically related organisms. This is intended to make the classification more stable in the long term and easier to update. Some of the main groups of protists, which may be treated as phyla, are listed in the taxobox, upper right.[35] Many are thought to be monophyletic, though there is still uncertainty. For instance, the excavates are probably not monophyletic and the chromalveolates are probably only monophyletic if the haptophytes and cryptomonads are excluded.[36]
Metabolism
Nutrition can vary according to the type of protist. Most eukaryotic algae are autotrophic, but the pigments were lost in some groups. Other protists are heterotrophic, and may present phagotrophy, osmotrophy, saprotrophy or parasitism. Some are mixotrophic. Some protists that do not have / lost chloroplasts/mitochondria have entered into endosymbiontic relationship with other bacteria/algae to replace the missing functionality. For ex. Paramecium bursaria & Paulinella have captured a green algae (Zoochlorella) & a cyanobacterium respectively that act as a replacement for chloroplast. Meanwhile, a protist, Mixotricha paradoxa that has lost its mitochondria uses endosymbiontic bacteria as mitochondria and ectosymbiontic hair-like bacteria (Treponema spirochetes) for locomotion.
Many protists are flagellate, for example, and filter feeding can take place where the flagella find prey. Other protists can engulf bacteria and other food particles, by extending their cell membrane around them to form a food vacuole and digest them internally, in a process termed phagocytosis.
| Nutritional type | Source of energy | Source of carbon | Examples |
|---|---|---|---|
| Photoautotrophs | Sunlight | Organic compounds or carbon fixation | Most algae |
| Chemoheterotrophs | Organic compounds | Organic compounds | Apicomplexa, Trypanosomes or Amoebae |
Reproduction
Some protists reproduce sexually using gametes, while others reproduce asexually by binary fission.
Some species, for example Plasmodium falciparum, have extremely complex life cycles that involve multiple forms of the organism, some of which reproduce sexually and others asexually.[37] However, it is unclear how frequently sexual reproduction causes genetic exchange between different strains of Plasmodium in nature and most populations of parasitic protists may be clonal lines that rarely exchange genes with other members of their species.[38]
Eukaryotes emerged in evolution more than 1.5 billion years ago.[39] The earliest eukaryotes were likely protists. Although sexual reproduction is widespread among extant eukaryotes, it seemed unlikely until recently, that sex could be a primordial and fundamental characteristic of eukaryotes. A principal reason for this view was that sex appeared to be lacking in certain pathogenic protists whose ancestors branched off early from the eukaryotic family tree. However, several of these protists are now known to be capable of, or to recently have had the capability for, meiosis and hence sexual reproduction. For example, the common intestinal parasite Giardia lamblia was once considered to be a descendant of a protist lineage that predated the emergence of meiosis and sex. However, G. lamblia was recently found to have a core set of genes that function in meiosis and that are widely present among sexual eukaryotes.[40] These results suggested that G. lamblia is capable of meiosis and thus sexual reproduction. Furthermore, direct evidence for meiotic recombination, indicative of sex, was also found in G. lamblia.[41]
The pathogenic parasitic protists of the genus Leishmania have been shown to be capable of a sexual cycle in the invertebrate vector, likened to the meiosis undertaken in the trypanosomes.[42]
Trichomonas vaginalis, a parasitic protist, is not known to undergo meiosis, but when Malik et al.[43] tested for 29 genes that function in meiosis, they found 27 to be present, including 8 of 9 genes specific to meiosis in model eukaryotes. These findings suggest that T. vaginalis may be capable of meiosis. Since 21 of the 29 meiotic genes were also present in G. lamblia, it appears that most of these meiotic genes were likely present in a common ancestor of T. vaginalis and G. lamblia. These two species are descendants of protist lineages that are highly divergent among eukaryotes, leading Malik et al.[43] to suggest that these meiotic genes were likely present in a common ancestor of all eukaryotes.
Based on a phylogenetic analysis, Dacks and Roger proposed that facultative sex was present in the common ancestor of all eukaryotes.[44]
This view was further supported by a study of amoebae by Lahr et al.[45] Amoeba have generally been regarded as asexual protists. However these authors describe evidence that most amoeboid lineages are anciently sexual, and that the majority of asexual groups likely arose recently and independently. It should be noted that early researchers (e.g., Calkins) have interpreted phenomena related to chromidia (chromatin granules free in the cytoplasm) in amoeboid organisms as sexual reproduction.[46]
Protists generally reproduce asexually under favorable environmental conditions, but tend to reproduce sexually under stressful conditions, such as starvation or heat shock.[47] Oxidative stress, which is associated with the production of reactive oxygen species leading to DNA damage, also appears to be an important factor in the induction of sex in protists.[47]
Ecology
Protists live in almost any environment that contains liquid water. Many protists, such as algae, are photosynthetic and are vital primary producers in ecosystems, particularly in the ocean as part of the plankton. Other protists include pathogenic species such as the kinetoplastid Trypanosoma brucei, which causes sleeping sickness and species of the apicomplexan Plasmodium which cause malaria.
Parasitism: role as pathogens
Some protists are significant parasites of animals (e.g., five species of the parasitic genus Plasmodium cause malaria in humans and many others cause similar diseases in other vertebrates), plants (the oomycete Phytophthora infestans causes late blight in potatoes)[48] or even of other protists.[49][50] Protist pathogens share many metabolic pathways with their eukaryotic hosts. This makes therapeutic target development extremely difficult – a drug that harms a protist parasite is also likely to harm its animal/plant host. A more thorough understanding of protist biology may allow these diseases to be treated more efficiently. For example, the apicoplast (a nonphotosynthetic chloroplast but essential to carry out important functions other than photosynthesis) present in apicomplexans provides an attractive target for treating diseases caused by dangerous pathogens such as plasmodium.
Recent papers have proposed the use of viruses to treat infections caused by protozoa.[51][52]
Researchers from the Agricultural Research Service are taking advantage of protists as pathogens to control red imported fire ant (Solenopsis invicta) populations in Argentina. Spore-producing protists such as Kneallhazia solenopsae (recognized as a sister clade or the closest relative to the fungus kingdom now)[53] can reduce red fire ant populations by 53–100%.[54] Researchers have also been able to infect phorid fly parasitoids of the ant with the protist without harming the flies. This turns the flies into a vector that can spreads the pathogenic protist between red fire ant colonies.[55]
Fossil record
Many protists have neither hard parts nor resistant spores, and their fossils are extremely rare or unknown. Examples of such groups include the apicomplexans,[56] most ciliates,[57] some green algae (the Klebsormidiales),[58] choanoflagellates,[59] oomycetes,[60] brown algae,[61] yellow-green algae,[62] excavates (e.g., euglenids).[63] Some of these have been found preserved in amber (fossilized tree resin) or under unusual conditions (e.g., Paleoleishmania, a kinetoplastid).
Others are relatively common in the fossil record,[64] as the diatoms,[65] golden algae,[66] haptophytes (coccoliths),[67] silicoflagellates, tintinnids (ciliates), dinoflagellates,[68] green algae,[69] red algae,[70] heliozoans, radiolarians,[71] foraminiferans,[72] ebriids and testate amoebae (euglyphids, arcellaceans).[73] Some are even used as paleoecological indicators to reconstruct ancient environments.
More probable eukaryote fossils begin to appear at about 1.8 billion years ago, the acritarchs, spherical fossils of likely algal protists.[74] Another possible representant of early fossil eukaryotes are the Gabonionta.
See also
Notes
- ↑ In the original 4-kingdom model proposed in 1959, Protista included all unicellular microorganisms such as bacteria. Herbert Copeland proposed separate kingdoms – Mychota – for prokaryotes and – Protoctista – for eukaryotes (including fungi) that were neither plants nor animals. Copeland's distinction between prokaryotic and eukaryotic cells was eventually critical in Whittaker proposing a final five-kingdom system, even though he resisted it for over a decade.[4][5]
References
- ↑ Adl Sina M.; et al. (2012). "The revised classification of eukaryotes" (PDF). Journal of Eukaryotic Microbiology. 59 (5): 429–514. doi:10.1111/j.1550-7408.2012.00644.x. PMC 3483872
 . PMID 23020233.
. PMID 23020233. - ↑ Whittaker, R. H. (1969). "New Concepts of Kingdoms of Organisms". Science. 163 (3863): 150–60. doi:10.1126/science.163.3863.150. PMID 5762760.
- ↑ "whittaker new concepts of kingdoms – Google Scholar". scholar.google.ca. Retrieved 2016-02-28.
- 1 2 Whittaker, R. H. (1959). "On the Broad Classification of Organisms". Quarterly Review of Biology. 34 (3): 210. doi:10.1086/402733. JSTOR 2816520.
- ↑ Hagen, Joel B. (2012). "depiction of Whittaker's early four-kingdom system, based on three modes of nutrition and the distinction between unicellular and multicellular body plans". BioScience. 62: 67. doi:10.1525/bio.2012.62.1.11.
- ↑ "Systematics of the Eukaryota". Retrieved 2009-05-31.
- ↑ Simonite T (November 2005). "Protists push animals aside in rule revamp". Nature. 438 (7064): 8–9. Bibcode:2005Natur.438....8S. doi:10.1038/438008b. PMID 16267517.
- ↑ Harper, David; Benton, Michael (2009). Introduction to Paleobiology and the Fossil Record. Wiley-Blackwell. p. 207. ISBN 1-4051-4157-3.
- ↑ Margulis, Lynn; Chapman, Michael J. (2009-03-19). Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth. Academic Press. ISBN 9780080920146.
- ↑ Siddiqui, Ruqaiyyah; Kulsoom, Huma; Lalani, Salima; Khan, Naveed Ahmed (2016-07-01). "Isolation of Balamuthia mandrillaris-specific antibody fragments from a bacteriophage antibody display library". Experimental Parasitology. 166: 94–96. doi:10.1016/j.exppara.2016.04.001.
- ↑ Corliss, J.O. (1995). "The ambiregnal protists and the codes of nomenclature: a brief review of the problem and of proposed solutions". Bulletin of Zoological Nomenclature. 52: 11–17.
- ↑ Barnes, Richard Stephen Kent (2001). The Invertebrates: A Synthesis. Wiley-Blackwell. p. 41. ISBN 978-0-632-04761-1.
- ↑ Ratcliff, Marc J. (2009). "The Emergence of the Systematics of Infusoria". In: The Quest for the Invisible: Microscopy in the Enlightenment. Aldershot: Ashgate.
- ↑ Sharma, O. P. (1986). Textbook of Algae. McGraw Hill. p. 22.
- ↑ Fauré-Frémiet, E. & Théodoridès, J. (1972). État des connaissances sur la structure des Protozoaires avant la formulation de la Théorie cellulaire. Revue d'histoire des sciences, 27–44.
- ↑ The Flagellates. Unity, diversity and evolution. Ed.: Barry S. C. Leadbeater and J. C. Green Taylor and Francis, London 2000, p. 3.
- 1 2 Scamardella, J. M. (1999). "Not plants or animals: a brief history of the origin of Kingdoms Protozoa, Protista and Protoctista" (PDF). International Microbiology. 2: 207–221.
- ↑ Rothschild LJ (1989). "Protozoa, Protista, Protoctista: what's in a name?". J Hist Biol. 22 (2): 277–305. doi:10.1007/BF00139515. PMID 11542176.
- ↑ Copeland, H. F. (1938). "The Kingdoms of Organisms". Quarterly Review of Biology. 13 (4): 383. doi:10.1086/394568. JSTOR 2808554.
- ↑ Whittaker RH (January 1969). "New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms". Science. 163 (3863): 150–60. Bibcode:1969Sci...163..150W. doi:10.1126/science.163.3863.150. PMID 5762760.
- ↑ Stechmann, Alexandra; Thomas Cavalier-Smith (2003). "The root of the eukaryote tree pinpointed" (PDF). Current Biology. 13 (17): R665–R666. doi:10.1016/S0960-9822(03)00602-X. PMID 12956967. Retrieved 15 May 2011.
- 1 2 O’Malley, M. A.; Simpson, A. G. B.; Roger, A. J. (2012). "The other eukaryotes in light of evolutionary protistology". Biology & Philosophy. 28 (2): 299–330. doi:10.1007/s10539-012-9354-y.
- ↑ Martin Schlegela, Norbert Hülsmannb (2 August 2007). "Protists – A textbook example for a paraphyletic taxon". Volume 7, Issue 2. Organisms Diversity & Evolution. pp. Pages 166–172. Retrieved 11 June 2016.
- ↑ "Protista". microbeworld.org. Retrieved 11 June 2016.
- ↑ Štolc, A. (1899). "Actinomyxidies, nouveau groupe de Mesozoaires parent des Myxosporidies". Bull. Int. L'Acad. Sci. Bohème. 12: 1–12.
- 1 2 Adl SM, Simpson AG, Farmer MA, et al. (2005). "The new higher level classification of eukaryotes with emphasis on the taxonomy of protists". J. Eukaryot. Microbiol. 52 (5): 399–451. doi:10.1111/j.1550-7408.2005.00053.x. PMID 16248873.
- ↑ Pitelka, D. R. (1963). Electron-Microscopic Structure of Protozoa. Pergamon Press, Oxford.
- ↑ Berner, T. (1993). Ultrastructure of Microalgae. Boca Raton: CRC Press. ISBN 0849363233
- ↑ Beckett, A., Heath, I. B., and Mclaughlin, D. J. (1974). An Atlas of Fungal Ultrastructure. Longman, Green, New York.
- ↑ Ragan M.A. & Chapman D.J. (1978). A Biochemical Phylogeny of the Protists. London, New York: Academic Press. ISBN 0323155618
- ↑ Lewin R. A. (1974). "Biochemical taxonomy", pp. 1–39 in Algal Physiology and Biochemistry, Stewart W. D. P. (ed.). Blackwell Scientific Publications, Oxford. ISBN 0520024109
- ↑ Oren, A., & Papke, R. T. (2010). Molecular phylogeny of microorganisms. Norfolk, UK: Caister Academic Press. ISBN 1904455670
- ↑ Horner, D. S., & Hirt, R. P. (2004). "An overview on eukaryote origins and evolution: the beauty of the cell and the fabulous gene phylogenies", pp. 1–26 in Hirt, R.P. & D.S. Horner. Organelles, Genomes and Eukaryote Phylogeny, An Evolutionary Synthesis in the Age of Genomics. New York: CRC Press. ISBN 0203508939
- ↑ Lahr, D. J. G.; Laughinghouse, H. D.; Oliverio, A. M.; Gao, F.; Katz, L. A. (2014). "How discordant morphological and molecular evolution among microorganisms can revise our notions of biodiversity on Earth". BioEssays. 36 (10): 950–9. doi:10.1002/bies.201400056. PMID 25156897.
- ↑ Cavalier-Smith T, Chao EE; Chao (October 2003). "Phylogeny and classification of phylum Cercozoa (Protozoa)". Protist. 154 (3–4): 341–58. doi:10.1078/143446103322454112. PMID 14658494.
- ↑ Laura Wegener Parfrey; Erika Barbero; Elyse Lasser; Micah Dunthorn; Debashish Bhattacharya; David J Patterson & Laura A Katz (December 2006). "Evaluating Support for the Current Classification of Eukaryotic Diversity". PLoS Genet. 2 (12): e220. doi:10.1371/journal.pgen.0020220. PMC 1713255. PMID 17194223.
- ↑ Talman AM, Domarle O, McKenzie FE, Ariey F, Robert V; Domarle; McKenzie; Ariey; Robert (July 2004). "Gametocytogenesis: the puberty of Plasmodium falciparum". Malar. J. 3: 24. doi:10.1186/1475-2875-3-24. PMC 497046. PMID 15253774.
- ↑ Tibayrenc M, Kjellberg F, Arnaud J; et al. (June 1991). "Are eukaryotic microorganisms clonal or sexual? A population genetics vantage". Proc. Natl. Acad. Sci. U.S.A. 88 (12): 5129–33. Bibcode:1991PNAS...88.5129T. doi:10.1073/pnas.88.12.5129. PMC 51825. PMID 1675793.
- ↑ Javaux EJ, Knoll AH, Walter MR; Knoll; Walter (July 2001). "Morphological and ecological complexity in early eukaryotic ecosystems". Nature. 412 (6842): 66–9. doi:10.1038/35083562. PMID 11452306.
- ↑ Ramesh MA, Malik SB, Logsdon JM; Malik; Logsdon Jr (January 2005). "A phylogenomic inventory of meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis". Curr. Biol. 15 (2): 185–91. doi:10.1016/j.cub.2005.01.003. PMID 15668177.
- ↑ Cooper MA, Adam RD, Worobey M, Sterling CR; Adam; Worobey; Sterling (November 2007). "Population genetics provides evidence for recombination in Giardia". Curr. Biol. 17 (22): 1984–8. doi:10.1016/j.cub.2007.10.020. PMID 17980591.
- ↑ Akopyants NS, Kimblin N, Secundino N; et al. (April 2009). "Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector". Science. 324 (5924): 265–8. Bibcode:2009Sci...324..265A. doi:10.1126/science.1169464. PMC 2729066. PMID 19359589.
- 1 2 Malik SB, Pightling AW, Stefaniak LM, Schurko AM, Logsdon JM; Pightling; Stefaniak; Schurko; Logsdon Jr (2008). Hahn, Matthew W, ed. "An expanded inventory of conserved meiotic genes provides evidence for sex in Trichomonas vaginalis". PLoS ONE. 3 (8): e2879. Bibcode:2008PLoSO...3.2879M. doi:10.1371/journal.pone.0002879. PMC 2488364. PMID 18663385.
- ↑ Dacks J, Roger AJ; Roger (1999). "The first sexual lineage and the relevance of facultative sex". J. Mol. Evol. 48 (6): 779–83. doi:10.1007/PL00013156. PMID 10229582.
- ↑ Lahr DJ, Parfrey LW, Mitchell EA, Katz LA, Lara E; Parfrey; Mitchell; Katz; Lara (July 2011). "The chastity of amoebae: re-evaluating evidence for sex in amoeboid organisms". Proc. Biol. Sci. 278 (1715): 2081–90. doi:10.1098/rspb.2011.0289. PMC 3107637. PMID 21429931.
- ↑ Dobell, C. (1909). "Chromidia and the binuclearity hypotheses: a review and a criticism" (PDF). Quarterly Journal of Microscopical Science. 53: 279–326.
- 1 2 Bernstein H, Bernstein C, Michod RE (2012). "DNA repair as the primary adaptive function of sex in bacteria and eukaryotes". Chapter 1: pp. 1–49 in DNA Repair: New Research, Sakura Kimura and Sora Shimizu (eds.). Nova Sci. Publ., Hauppauge, N.Y. ISBN 978-1-62100-808-8
- ↑ Campbell, N. and Reese, J. (2008) Biology. Pearson Benjamin Cummings; 8 ed. ISBN 0805368442. pp. 583, 588
- ↑ Lauckner, G. (1980). Diseases of protozoa. In: Diseases of Marine Animals. Kinne, O. (ed.). Vol. 1, p. 84, John Wiley & Sons, Chichester, UK.
- ↑ Cox, F.E.G. (1991). Systematics of parasitic protozoa. In: Kreier, J.P. & J. R. Baker (ed.). Parasitic Protozoa, 2nd ed., vol. 1. San Diego: Academic Press.
- ↑ Keen, E. C. (2013). "Beyond phage therapy: Virotherapy of protozoal diseases". Future Microbiology. 8 (7): 821–823. doi:10.2217/FMB.13.48. PMID 23841627.
- ↑ Hyman, P.; Atterbury, R.; Barrow, P. (2013). "Fleas and smaller fleas: Virotherapy for parasite infections". Trends in Microbiology. 21 (5): 215–220. doi:10.1016/j.tim.2013.02.006. PMID 23540830.
- ↑ Liu, Y. J.; Hodson, M. C.; Hall, B. D. (2006). "phylogenetic structure of Kingdom Fungi inferred from RNA polymerase II subunit genes". BMC Evolutionary Biology. 6: 74. doi:10.1186/1471-2148-6-74. PMC 1599754. PMID 17010206.
- ↑ "ARS Parasite Collections Assist Research and Diagnoses". USDA Agricultural Research Service. January 28, 2010.
- ↑ Durham, Sharon (January 28, 2010) ARS Parasite Collections Assist Research and Diagnoses. Ars.usda.gov. Retrieved 2014-03-20.
- ↑ Introduction to the Apicomplexa. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of the Ciliata. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Klebsormidiales. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Choanoflagellata. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Oomycota. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Phaeophyta. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Xanthophyta. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Basal Eukaryotes. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Why Is The Museum On The Web?. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of Diatoms. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Chrysophyta. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Prymnesiophyta. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of the Dinoflagellata. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Systematics of the "Green Algae", Part 1. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of the Rhodophyta. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of the Radiolaria. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of Foraminifera. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Testaceafilosea. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of the Eukaryota. Ucmp.berkeley.edu. Retrieved 2014-03-20.
Bibliography
General
- Haeckel, E. Das Protistenreich. Leipzig, 1878.
- Hausmann, K., N. Hulsmann, R. Radek. Protistology. Schweizerbart'sche Verlagsbuchshandlung, Stuttgart, 2003.
- Margulis, L., J.O. Corliss, M. Melkonian, D.J. Chapman. Handbook of Protoctista. Jones and Bartlett Publishers, Boston, 1990.
- Margulis, L., K.V. Schwartz. Five Kingdoms: An Illustrated Guide to the Phyla of Life on Earth, 3rd ed. New York: W.H. Freeman, 1998.
- Margulis, L., L. Olendzenski, H.I. McKhann. Illustrated Glossary of the Protoctista, 1993.
- Margulis, L., M.J. Chapman. Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth. Amsterdam: Academic Press/Elsevier, 2009.
- Schaechter, M. Eukaryotic microbes. Amsterdam, Academic Press, 2012.
Physiology, ecology and paleontology
- Foissner, W.; D.L. Hawksworth. Protist Diversity and Geographical Distribution. Dordrecht: Springer, 2009
- Fontaneto, D. Biogeography of Microscopic Organisms. Is Everything Small Everywhere? Cambridge University Press, Cambridge, 2011.
- Levandowsky, M. Physiological Adaptations of Protists. In: Cell physiology sourcebook : essentials of membrane biophysics. Amsterdam; Boston: Elsevier/AP, 2012.
- Moore, R. C., and other editors. Treatise on Invertebrate Paleontology. Protista, part B (vol. 1, Charophyta, vol. 2, Chrysomonadida, Coccolithophorida, Charophyta, Diatomacea & Pyrrhophyta), part C (Sarcodina, Chiefly "Thecamoebians" and Foraminiferida) and part D (Chiefly Radiolaria and Tintinnina). Boulder, Colorado: Geological Society of America; & Lawrence, Kansas: University of Kansas Press.
External links
| Wikispecies has information related to: Protista |
| Wikispecies has information related to: Protoctista |
| Wikimedia Commons has media related to Protista. |
- Tree of Life: Eukaryotes
- A java applet for exploring the new higher level classification of eukaryotes
- Plankton Chronicles – Protists – Cells in the Sea – video
- Holt, Jack R. and Carlos A. Iudica. (2013). Diversity of Life. http://comenius.susqu.edu/biol/202/Taxa.htm. Last modified: 11/18/13.
- Tsukii, Y. (1996). Protist Information Server (database of protist images). Laboratory of Biology, Hosei University.. Updated: March 22, 2016.