Ribozyme
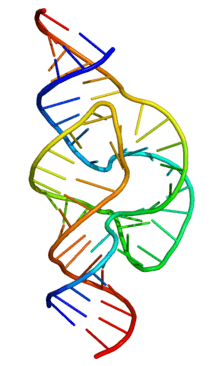
Ribozymes (ribonucleic acid enzymes) are RNA molecules that are capable of catalyzing specific biochemical reactions, similar to the action of protein enzymes. The 1982 discovery of ribozymes demonstrated that RNA can be both genetic material (like DNA) and a biological catalyst (like protein enzymes), and contributed to the RNA world hypothesis, which suggests that RNA may have been important in the evolution of prebiotic self-replicating systems. The most common activities of natural or in vitro-evolved ribozymes are the cleavage or ligation of RNA and DNA and peptide bond formation.[1] Within the ribosome, ribozymes function as part of the large subunit ribosomal RNA to link amino acids during protein synthesis. They also participate in a variety of RNA processing reactions, including RNA splicing, viral replication, and transfer RNA biosynthesis. Examples of ribozymes include the hammerhead ribozyme, the VS ribozyme, Leadzyme and the hairpin ribozyme.
Investigators studying the origin of life have produced ribozymes in the laboratory that are capable of catalyzing their own synthesis from activated monomers under very specific conditions, such as an RNA polymerase ribozyme.[2] Mutagenesis and selection has been performed resulting in isolation of improved variants of the "Round-18" polymerase ribozyme from 2001. "B6.61" is able to add up to 20 nucleotides to a primer template in 24 hours, until it decomposes by cleavage of its phosphodiester bonds.[3] The "tC19Z" ribozyme can add up to 95 nucleotides with a fidelity of 0.0083 mutations/nucleotide.[4]
Attempts have been made to develop ribozymes as therapeutic agents, as enzymes which target defined RNA sequences for cleavage, as biosensors, and for applications in functional genomics and gene discovery.[5]
Discovery

Before the discovery of ribozymes, enzymes, which are defined as catalytic proteins,[6] were the only known biological catalysts. In 1967, Carl Woese, Francis Crick, and Leslie Orgel were the first to suggest that RNA could act as a catalyst. This idea was based upon the discovery that RNA can form complex secondary structures.[7] Ribozymes were discovered in the early 1980s by Thomas R. Cech, who was studying RNA splicing in the ciliated protozoan Tetrahymena thermophila and by Sidney Altman and Norman Pace, who were studying the bacterial RNase P complex. These ribozymes were found in the intron of an RNA transcript, which removed itself from the transcript, as well as in the RNA component of the RNase P complex, which is involved in the maturation of pre-tRNAs. In 1989, Thomas R. Cech and Sidney Altman shared the Nobel Prize in chemistry for their "discovery of catalytic properties of RNA."[8] The term ribozyme was first introduced by Kelly Kruger et al. in 1982 in a paper published in Cell.[9]
It had been a firmly established belief in biology that catalysis was reserved for proteins. However, the idea of RNA catalysis is motivated in part by the old question regarding the origin of life: Which comes first, enzymes that do the work of the cell or nucleic acids that carry the information required to produce the enzymes? The concept of "ribonucleic acids as catalysts" circumvents this problem. RNA, in essence, can be both the chicken and the egg.[10]
In the 1980s Thomas Cech, at the University of Colorado at Boulder, was studying the excision of introns in a ribosomal RNA gene in Tetrahymena thermophila. While trying to purify the enzyme responsible for the splicing reaction, he found that the intron could be spliced out in the absence of any added cell extract. As much as they tried, Cech and his colleagues could not identify any protein associated with the splicing reaction. After much work, Cech proposed that the intron sequence portion of the RNA could break and reform phosphodiester bonds. At about the same time, Sidney Altman, a professor at Yale University, was studying the way tRNA molecules are processed in the cell when he and his colleagues isolated an enzyme called RNase-P, which is responsible for conversion of a precursor tRNA into the active tRNA. Much to their surprise, they found that RNase-P contained RNA in addition to protein and that RNA was an essential component of the active enzyme. This was such a foreign idea that they had difficulty publishing their findings. The following year, Altman demonstrated that RNA can act as a catalyst by showing that the RNase-P RNA subunit could catalyze the cleavage of precursor tRNA into active tRNA in the absence of any protein component.
Since Cech's and Altman's discovery, other investigators have discovered other examples of self-cleaving RNA or catalytic RNA molecules. Many ribozymes have either a hairpin – or hammerhead – shaped active center and a unique secondary structure that allows them to cleave other RNA molecules at specific sequences. It is now possible to make ribozymes that will specifically cleave any RNA molecule. These RNA catalysts may have pharmaceutical applications. For example, a ribozyme has been designed to cleave the RNA of HIV. If such a ribozyme were made by a cell, all incoming virus particles would have their RNA genome cleaved by the ribozyme, which would prevent infection.
Structure and mechanism
Despite having only four choices for each monomer unit (nucleotides), compared to 20 amino acid side chains found in proteins, ribozymes have diverse structures and mechanisms. In many cases they are able to mimic the mechanism used by their protein counterparts. For example, in self cleaving ribozyme RNAs, an in-line SN2 reaction is carried out using the 2’ hydroxyl group as a nucleophile attacking the bridging phosphate and causing 5’ oxygen of the N+1 base to act as a leaving group. In comparison, RNase A, a protein that catalyzes the same reaction, uses a coordinating histidine and lysine to act as a base to attack the phosphate backbone.[1]
Like many protein enzymes metal binding is also critical to the function of many ribozymes.[11] Often these interactions use both the phosphate backbone and the base of the nucleotide, causing drastic conformational changes.[12]
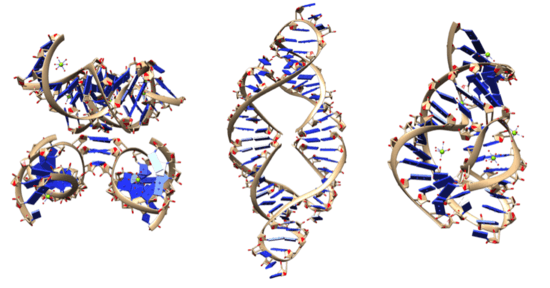
Activity
Although most ribozymes are quite rare in the cell, their roles are sometimes essential to life. For example, the functional part of the ribosome, the molecular machine that translates RNA into proteins, is fundamentally a ribozyme, composed of RNA tertiary structural motifs that are often coordinated to metal ions such as Mg2+ as cofactors. In a model system, there is no requirement for divalent cations in a five-nucleotide RNA catalyzing trans-phenylalanation of a four-nucleotide substrate with 3 base pairs complementary with the catalyst, where the catalyst/substrate were devised by truncation of the C3 ribozyme.[13] RNA may catalyze folding of the pathological protein conformation of a prion in a manner similar to that of a chaperonin,[14] and may be involved in the viral concatemer cleavage that precedes the packing of viral genetic material into some virons.
RNA can also act as a hereditary molecule, which encouraged Walter Gilbert to propose that in the distant past, the cell used RNA as both the genetic material and the structural and catalytic molecule rather than dividing these functions between DNA and protein as they are today; this hypothesis is known as the "RNA world hypothesis" of the origin of life.[15] Evidence that ribozymes were the first molecular machines used by early life suggests that they are in effect "molecular fossils".
Artificial ribozymes
Since the discovery of ribozymes that exist in living organisms, there has been interest in the study of new synthetic ribozymes made in the laboratory. For example, artificially-produced self-cleaving RNAs that have good enzymatic activity have been produced. Tang and Breaker[16] isolated self-cleaving RNAs by in vitro selection of RNAs originating from random-sequence RNAs. Some of the synthetic ribozymes that were produced had novel structures, while some were similar to the naturally occurring hammerhead ribozyme. In 2015, researchers at Northwestern University and the University of Illinois at Chicago have engineered a tethered ribosome that works nearly as well as the authentic cellular component that produces all the proteins and enzymes within the cell. Called Ribosome-T, or Ribo-T, the artificial ribosome was created by Michael Jewett and Alexander Mankin.[17] The techniques used to create artificial ribozymes involve directed evolution. This approach takes advantage of RNA's dual nature as both a catalyst and an informational polymer, making it easy for an investigator to produce vast populations of RNA catalysts using polymerase enzymes. The ribozymes are mutated by reverse transcribing them with reverse transcriptase into various cDNA and amplified with error-prone PCR. The selection parameters in these experiments often differ. One approach for selecting a ligase ribozyme involves using biotin tags, which are covalently linked to the substrate. If a molecule possesses the desired ligase activity, a streptavidin matrix can be used to recover the active molecules.
Lincoln and Joyce developed an RNA enzyme system capable of self replication in about an hour. By utilizing molecular competition (in vitro evolution) of a candidate RNAmixture, a pair of ribozymes emerged, in which each synthesizes the other by joining synthetic oligonucleotides, with no protein present.[18]
Although not true catalysts, the creation of artificial self-cleaving riboswitches, termed aptazymes, has also been an active area of research. Riboswitches are regulatory RNA motifs that change their structure in response to a small molecule ligand to regulate translation. While there are many known natural riboswitches that bind a wide array of metabolites and other small organic molecules, only one ribozyme based on a riboswitch has been described, glmS.[19] Early work in characterizing self-cleaving riboswitches was focused on using theophylline as the ligand. In these studies an RNA hairpin is formed which blocks the ribosome binding site, thus inhibiting translation. In the presence of the ligand, in these cases theophylline, the regulatory RNA region is cleaved off, allowing the ribosome to bind and translate the target gene. Much of this RNA engineering work was based on rational design and previously determined RNA structures rather than directed evolution as in the above examples. More recent work has broadened the ligands used in ribozyme riboswitches to include thymine pyrophosphate (2). Fluorescence-activated_cell_sorting has also been used to engineering aptazymes.[20]
Applications
Ribozymes have been proposed and developed for the treatment of disease through gene therapy (3). One major challenge of using RNA based enzymes as a therapeutic is the short half-life of the catalytic RNA molecules in the body. To combat this, the 2’ position on the ribose is modified to improve RNA stability. One area of ribozyme gene therapy has been the inhibition of RNA-based viruses.
A type of synthetic ribozyme directed against HIV RNA called gene shears has been developed and has entered clinical testing for HIV infection.[21][22]
Similarly, a ribozyme has been designed to target the hepatitis C virus RNA. The ribozyme is able to cleave the conserved regions of the virus’s genome which has been shown to reduce the virus in mammalian cell culture.[23] Despite these efforts by researchers, these projects have remained in the preclinical stage.
Known ribozymes
Well validated naturally occurring ribozyme classes:
- GIR1 branching ribozyme[24]
- glmS ribozyme
- Group I self-splicing intron
- Group II self-splicing intron - Spliceosome is likely derived from Group II self-splicing ribozymes.[25]
- Hairpin ribozyme
- Hammerhead ribozyme
- HDV ribozyme
- rRNA - Found in all living cells and links amino acids to form proteins.
- RNase P
- Twister ribozyme
- Twister sister ribozyme
- VS ribozyme
- Pistol ribozyme
- Hatchet ribozyme
See also
Further reading
- Astrid Sigel, Helmut Sigel and Roland K. O. Sigel, ed. (2011). Structural and catalytic roles of metal ions in RNA. Metal Ion in Life Sciences. 9. Cambridge, U.K.: RSC Publishing. doi:10.1039/9781849732512. ISBN 978-1-84973-251-2.
- Johnson-Buck, Alexander E.; McDowell, Sarah E.; Walter, Nils. G. "Chapter 6. Metal Ions: Supporting Actors in the Playbook of Small Ribozymes". pp. 175–196. doi:10.1039/9781849732512-00175. Missing or empty
|title=(help) - Donghi, Daniela; Schnabl, Joachim. "Chapter 7. Multiple roles of metal ions in large ribozymes". pp. 197–234. doi:10.1039/9781849732512-00197. Missing or empty
|title=(help) - Trappl, Krista; Polacek, Norbert. "Chapter 9. The Ribosome: A Molecular Machine Powered by RNA". pp. 253–275. doi:10.1039/9781849732512-00253. Missing or empty
|title=(help) - Suga, Hiroaki; Futai, Kazuki; Jin, Koichiro. "Chapter 10. Metal Ion Requirements in Artificial Ribozymes that Catalyze Aminoacylation". pp. 277–297. doi:10.1039/9781849732512-00277. Missing or empty
|title=(help) - Wedekind, Joseph E. "Chapter 11. Metal Ion Binding and Function in Natural and Artificial Small RNA Enzymes". pp. 299–345. doi:10.1039/9781849732512-00299. Missing or empty
|title=(help)
- Johnson-Buck, Alexander E.; McDowell, Sarah E.; Walter, Nils. G. "Chapter 6. Metal Ions: Supporting Actors in the Playbook of Small Ribozymes". pp. 175–196. doi:10.1039/9781849732512-00175. Missing or empty
- Doherty EA, Doudna JA (2001). "Ribozyme structures and mechanisms". Annu Rev Biophys Biomol Struct. 30: 457–475. doi:10.1146/annurev.biophys.30.1.457. PMID 11441810.
- Joyce GF (2004). "Directed evolution of nucleic acid enzymes". Annu. Rev. Biochem. 73: 791–836. doi:10.1146/annurev.biochem.73.011303.073717. PMID 15189159.
- Ikawa Y, Tsuda K, Matsumura S, Inoue T (September 2004). "De novo synthesis and development of an RNA enzyme". Proc. Natl. Acad. Sci. U.S.A. 101 (38): 13750–13755. Bibcode:2004PNAS..10113750I. doi:10.1073/pnas.0405886101. PMC 518828
 . PMID 15365187.
. PMID 15365187.
External links
Notes and references
- 1 2 Fedor M. J.; Williamson J. R. (2005). "The catalytic diversity of RNAs". Nat. Rev. Mol. Cell Biol. 6: 399–412. doi:10.1038/nrm1647.
- ↑ Johnston WK, Unrau PJ, Lawrence MS, Glasner ME, Bartel DP (May 2001). "RNA-catalyzed RNA polymerization: accurate and general RNA-templated primer extension". Science. 292 (5520): 1319–1325. Bibcode:2001Sci...292.1319J. doi:10.1126/science.1060786. PMID 11358999.
- ↑ Zaher HS, Unrau PJ (July 2007). "Selection of an improved RNA polymerase ribozyme with superior extension and fidelity". RNA. 13 (7): 1017–1026. doi:10.1261/rna.548807. PMC 1894930. PMID 17586759.
- ↑ Wochner A, Attwater J, Coulson A, Holliger P (April 2011). "Ribozyme-catalyzed transcription of an active ribozyme". Science. 332 (6026): 209–212. Bibcode:2011Sci...332..209W. doi:10.1126/science.1200752. PMID 21474753.
- ↑ Hean J, Weinberg MS (2008). "The Hammerhead Ribozyme Revisited: New Biological Insights for the Development of Therapeutic Agents and for Reverse Genomics Applications". In Morris KL. RNA and the Regulation of Gene Expression: A Hidden Layer of Complexity. Norfolk, England: Caister Academic Press. ISBN 1-904455-25-5.
- ↑ Enzyme definition Dictionary.com Accessed 6 April 2007
- ↑ Carl Woese, The Genetic Code (New York: Harper and Row, 1967).
- ↑ The Nobel Prize in Chemistry 1989 was awarded to Thomas R. Cech and Sidney Altman "for their discovery of catalytic properties of RNA".
- ↑ Kruger K, Grabowski PJ, Zaug AJ, Sands J, Gottschling DE, Cech TR (November 1982). "Self-splicing RNA: autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena". Cell. 31 (1): 147–57. doi:10.1016/0092-8674(82)90414-7. PMID 6297745.
- ↑ Visser CM (1984). "Evolution of biocatalysis 1. Possible pre-genetic-code RNA catalysts which are their own replicase". Orig. Life. 14 (1–4): 291–300. Bibcode:1984OrLi...14..291V. doi:10.1007/BF00933670. PMID 6205343.
- ↑ Pyle A. M. (1993). "Ribozymes: a distinct class of metalloenzymes". Sci. 261: 709–714. doi:10.1126/science.7688142. PMID 7688142.
- ↑ Freisinger E.; Sigel R. K. O. (2007). "From nucleotides to ribozymes—A comparison of their metal ion binding properties". Coord. Chem. Rev. 251: 1834–1851. doi:10.1016/j.ccr.2007.03.008.
- ↑ Turk RM, Chumachenko NV, Yarus M (March 2010). "Multiple translational products from a five-nucleotide ribozyme". Proc. Natl. Acad. Sci. U.S.A. 107 (10): 4585–4589. Bibcode:2010PNAS..107.4585T. doi:10.1073/pnas.0912895107. PMC 2826339. PMID 20176971.
- ↑ Supattapone S (June 2004). "Prion protein conversion in vitro". J. Mol. Med. 82 (6): 348–356. doi:10.1007/s00109-004-0534-3. PMID 15014886.
- ↑ Gilbert W (1986). "Origin of life: The RNA world". Nature. 319 (6055): 618. doi:10.1038/319618a0.
- ↑ Tang J, Breaker RR (May 2000). "Structural diversity of self-cleaving ribozymes". Proc. Natl. Acad. Sci. U.S.A. 97 (11): 5784–5789. Bibcode:2000PNAS...97.5784T. doi:10.1073/pnas.97.11.5784. PMC 18511. PMID 10823936.
- ↑ Engineer and Biologist Design First Artificial Ribosome - Designer ribosome could lead to new drugs and next-generation biomaterials published on July 31, 2015 by Northwestern University
- ↑ Lincoln TA, Joyce GF (February 2009). "Self-sustained replication of an RNA enzyme". Science. 323 (5918): 1229–1232. Bibcode:2009Sci...323.1229L. doi:10.1126/science.1167856. PMC 2652413. PMID 19131595.
- ↑ Winkler, W. C., Nahvi, A., Roth, A., Collins, J. A. & Breaker, R. R. Control of gene expression by a natural metabolite-responsive ribozyme. 4–9
- ↑ Lynch , Gallivan J. P. (2009). "A flow cytometry-based screen for synthetic riboswitches". Nucleic Acids Res. 37: 184–92. doi:10.1093/nar/gkn924.
- ↑ de Feyter R, Li P (June 2000). "Technology evaluation: HIV ribozyme gene therapy, Gene Shears Pty Ltd". Curr. Opin. Mol. Ther. 2 (3): 332–5. PMID 11249628.
- ↑ Khan AU (May 2006). "Ribozyme: a clinical tool". Clin. Chim. Acta. 367 (1–2): 20–27. doi:10.1016/j.cca.2005.11.023. PMID 16426595.
- ↑ He, C., Polyak, S. J., Gretch, D. R., Barr, D. & Kay, M. A. Elimination of Hepatitis C Virus RNA in Infected Human Hepatocytes by Adenovirus-Mediated Expression of Ribozymes. 70, 8782–8791 (1996).
- ↑ Nielsen H, Westhof E, Johansen S (2005). "An mRNA is capped by a 2', 5' lariat catalyzed by a group I-like ribozyme". Science. 309 (5740): 1584–1587. Bibcode:2005Sci...309.1584N. doi:10.1126/science.1113645. PMID 16141078.
- ↑ Fica SM, Tuttle N, Novak T, Li NS, Lu J, Koodathingal P, Dai K, Staley JP, Piccirilli JA (2013). "RNA catalyses nucelar pre-mRNA splicing". Nature. 503 (7475): 229–234. doi:10.1038/nature12734. PMID 24196718.