Mitochondrial Eve
| Haplogroup Modern humans | |
|---|---|
 | |
| Possible time of origin | c. 100–230 kya[1] |
| Possible place of origin | East Africa |
| Ancestor | n/a |
| Descendants | Mitochondrial macro-haplogroups L0, L1, and L5 |
| Defining mutations | None |
In human genetics, the Mitochondrial Eve (also mt-Eve, mt-MRCA) is the matrilineal most recent common ancestor (MRCA) of all currently living humans, i.e., the most recent woman from whom all living humans descend in an unbroken line purely through their mothers, and through the mothers of those mothers, back until all lines converge on one woman. Mitochondrial Eve lived later than Homo heidelbergensis and the emergence of Homo neanderthalensis, but earlier than the out of Africa migration,[2] but her age is not known with certainty; a 2009 estimate cites an age between c. 152 and 234 thousand years ago (95% CI);[3] a 2013 study cites a range of 99–148 thousand years ago.[4]
Because all mitochondrial DNA (mtDNA) generally (but see paternal mtDNA transmission) is passed from mother to offspring without recombination, all mtDNA in every living person is directly descended from hers by definition, differing only by the mutations that over generations have occurred in the germ cell mtDNA since the conception of the original "Mitochondrial Eve".
The male analog to the Mitochondrial Eve is the Y-chromosomal Adam, the member of Homo sapiens sapiens from whom all living humans are descended patrilineally. The inherited DNA in the male case is his nuclear Y chromosome rather than the mtDNA. Mitochondrial Eve and Y-chromosomal Adam need not have lived at the same time.[5]
As of 2013, estimates for mt-MRCA and Y-MRCA alike are still subject to substantial uncertainty; thus, Y-MRCA has been estimated to have lived during a wide range of times from 180,000 to 581,000 years ago[6][7][8] (with a most likely age of between 120,000 and 156,000 years ago, roughly consistent with the estimate for mt-MRCA[4][9] )
The name "Mitochondrial Eve" alludes to biblical Eve.[10] This has led to repeated misrepresentations or misconceptions in journalistic accounts on the topic. The title of "Mitochondrial Eve" is not permanently fixed to a single individual, but rather shifts forward in time over the course of human history as maternal lineages become extinct. Unlike her biblical namesake, she was not the only living human female of her time. However, by the definition of Mitochondrial Eve, her female contemporaries, though they may have descendants alive today, do not have any descendants today who descend in an unbroken female line of descent.
History
Molecular clock
Allan Wilson, of the University of California, Berkeley, with doctoral students Mark Stoneking and Rebecca L. Cann, worked on developing methods using a molecular clock in the late 1970s. Wesley M. Brown joined the team as a post-doc in 1978. In 1979 they found that mutation in mtDNA was unexpectedly fast, exhibiting 0.02 substitution per base (1%) in a million years, which means 5-10 times faster than in nuclear DNA, forming an exception in humans when compared to those of other primates including Guinea baboon, rhesus macaque, and guenon. This was the first crucial step in understanding the usefulness of mtDNA in estimating evolutionary divergence and relationships,[11] and they soon published a paper describing the evolutionary relationships among gorillas, chimpanzees (common chimpanzee and bonobo) and humans.[12]
Strictly on the evolution of modern humans, Brown studied mitochondrial fractions from 21 individuals (placentas from labour and delivery) of different ethnicity and geographic origin. In 1980, he discovered remarkable variations among humans, and by estimating the rate of mutation, published the first estimate on the age of the mt-MRCA, at 180,000 years ago.[13] But the shortcomings were that medical records were disassociated and he didn't know about the actual donors' ethnic backgrounds.
The study was advanced by doctoral student Rebecca L. Cann, who started collecting mtDNA in 1979 based on ethnicity. She collected samples from US hospitals from women (their placenta, to be precise) of diverse ethnic backgrounds, such as from Asia, South Pacific, Europe and Americans of African descent (she could not get samples from aboriginal Africans). Their initial observation of the mitochondrial genome of 100 individuals led them to resolve that it was possible to trace back individual female lineages back hundreds of generations thereby reaching the point of a common ancestor.[14] However, in 1983, Cann and Wilson failed to show common ancestry from 112 individuals of different ethnic groups, and they had no indication of African samples as the oldest or the lineage of the common ancestor using an analysis called Fitch-Margoliash trees.[15]
Even in Cann's thesis in 1982, in which more elaborate estimates (including PAUP and Neighbor joining in addition to Fitch-Margoliash trees) were used, African origin of all humankind was not yet convincing, especially to Wilson, though the hunch was evident there.[16] Prudence in Wilson was in part due to a publication of an eminent geneticist, Luigi Luca Cavalli-Sforza and his team in 1981 in which they concluded that mtDNA pointed most likely to human origin from Asia.[17]
In 1981, graduate student Mark Stoneking joined Wilson's lab for his PhD and immediately pursued Cann's work in collecting samples from aboriginal Australians and New Guineans from hospitals in Australia and New Guinea.
1987 publication
By 1985, data from the mtDNA of 145 women of different populations, and of two cell lines, HeLa and GM 3043, derived from a Black American and a !Kung respectively, was available. After more than 40 revisions of the draft, the manuscript was submitted to Nature in late 1985 or early 1986[16] and published on 1 January 1987. The published conclusion was that all current human mtDNA originated from a single population from Africa, at the time dated to 140,000 and 200,000 years ago.[18]
The dating for "Eve" was a blow to the multiregional hypothesis, which was being controversially discussed at the time, and a boost to the theory of the recent origin model.[19]
Cann, Stoneking and Wilson did not use the term "Mitochondrial Eve" or even the name "Eve" in their original paper; it appears to originate with a 1987 article in Science by Roger Lewin, headlined "The Unmasking of Mitochondrial Eve."[20] The biblical connotation was very clear from the start. The accompanying research news in Nature had the title "Out of the garden of Eden."[21] Wilson himself preferred the term "Lucky Mother" [22] and thought the use of the name Eve "regrettable."[20][23] But the concept of Eve caught on with the public and was repeated in a Newsweek cover story (11 January 1988 issue featured a depiction of Adam and Eve on the cover, with the title "The Search for Adam and Eve"),[24] and a cover story in Time on 26 January 1987.[25]
Criticism and new evidence
Shortly after the 1987 publication, criticism of its methodology and secondary conclusions was published.[26] Both the dating of mt-Eve and the relevance of the age of the purely matrilineal descent for population replacement was controversially discussed during the 1990s;[27][28][29][30] Alan Templeton (1997) asserted that the study did "not support the hypothesis of a recent African origin for all of humanity following a split between Africans and non-Africans 100,000 years ago" and also did "not support the hypothesis of a recent global replacement of humans coming out of Africa."[31]
Although the original research did have analytical limitations, the estimate on the age of the mt-MRCA has proven robust.[32][33] More recent age estimates have remained consistent with the 140–200 kya estimate published in 1987: A 2013 estimate dated Mitochondrial Eve to about 160 kya (within the reserved estimate of the original research) and Out of Africa II to about 95 kya.[34] Another 2013 study (based on genome sequencing of 69 people from 9 different populations) reported the age of Mitochondrial Eve between 99 to 148 kya and that of the Y-MRCA between 120 and 156 kya.[4]
Female and mitochondrial ancestry


Without a DNA sample, it is not possible to reconstruct the complete genetic makeup (genome) of any individual who died very long ago. By analysing descendants' DNA, however, parts of ancestral genomes are estimated by scientists. Mitochondrial DNA (mtDNA) and Y-chromosome DNA are commonly used to trace ancestry in this manner. mtDNA is generally passed un-mixed from mothers to children of both sexes, along the maternal line, or matrilineally.[35][36] Matrilineal descent goes back to our mothers, to their mothers, until all female lineages converge.
Branches are identified by one or more unique markers which give a mitochondrial "DNA signature" or "haplotype" (e.g. the CRS is a haplotype). Each marker is a DNA base-pair that has resulted from an SNP mutation. Scientists sort mitochondrial DNA results into more or less related groups, with more or less recent common ancestors. This leads to the construction of a DNA family tree where the branches are in biological terms clades, and the common ancestors such as Mitochondrial Eve sit at branching points in this tree. Major branches are said to define a haplogroup (e.g. CRS belongs to haplogroup H), and large branches containing several haplogroups are called "macro-haplogroups".
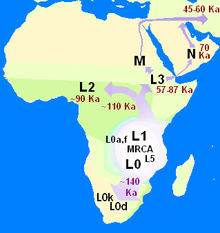
The mitochondrial clade which Mitochondrial Eve defines is the species Homo sapiens sapiens itself, or at least the current population or "chronospecies" as it exists today. In principle, earlier Eves can also be defined going beyond the species, for example one who is ancestral to both modern humanity and Neanderthals, or, further back, an "Eve" ancestral to all members of genus Homo and chimpanzees in genus Pan. According to current nomenclature, Mitochondrial Eve's haplogroup was within mitochondrial haplogroup L because this macro-haplogroup contains all surviving human mitochondrial lineages today, and she must predate the emergence of L0.
The variation of mitochondrial DNA between different people can be used to estimate the time back to a common ancestor, such as Mitochondrial Eve. This works because, along any particular line of descent, mitochondrial DNA accumulates mutations at the rate of approximately one every 3,500 years per nucleotide.[3][37][38] A certain number of these new variants will survive into modern times and be identifiable as distinct lineages. At the same time some branches, including even very old ones, come to an end, when the last family in a distinct branch has no daughters.
Mitochondrial Eve is the most recent common matrilineal ancestor for all modern humans. Whenever one of the two most ancient branch lines dies out, the MRCA will move to a more recent female ancestor, always the most recent mother to have more than one daughter with living maternal line descendants alive today. The number of mutations that can be found distinguishing modern people is determined by two criteria: firstly and most obviously, the time back to her, but secondly and less obviously by the varying rates at which new branches have come into existence and old branches have become extinct. By looking at the number of mutations which have been accumulated in different branches of this family tree, and looking at which geographical regions have the widest range of least related branches, the region where Eve lived can be proposed.
Common misconceptions
Not the only woman
One common misconception surrounding mitochondrial Eve is that since all women alive today descended in a direct unbroken female line from her, she must have been the only woman alive at the time.[5][39] However, nuclear DNA studies indicate that the size of the ancient human population never dropped below tens of thousands. Other women living during Eve's time have descendants alive today but not in a direct female line.
Not a fixed individual over time
The definition of mitochondrial Eve is fixed, but the woman in prehistory who fits this definition can change. That is, not only can our knowledge of when and where Mitochondrial Eve lived change due to new discoveries, but the actual mitochondrial Eve can change. The mitochondrial Eve can change, when a mother-daughter line comes to an end by chance. It follows from the definition of Mitochondrial Eve that she had at least two daughters who both have unbroken female lineages that have survived to the present day. In every generation mitochondrial lineages end – when a woman with unique mtDNA dies with no daughters. When the mitochondrial lineages of daughters of mitochondrial Eve die out, then the title of "Mitochondrial Eve" shifts forward from the remaining daughter through her matrilineal descendants, until the first descendant is reached who had two or more daughters who together have all living humans as their matrilineal descendants. Once a lineage has died out it is irretrievably lost and this mechanism can thus only shift the title of "Mitochondrial Eve" forward in time.
Because mtDNA mapping of humans is very incomplete, the discovery of living mtDNA lines which predate our current concept of "Mitochondrial Eve" could result in the title moving to an earlier woman. This happened to her male counterpart, "Y-chromosomal Adam," when older Y lines from Africa were discovered.
Not necessarily a contemporary of "Y-chromosomal Adam"
Sometimes mitochondrial Eve is assumed to have lived at the same time as Y-chromosomal Adam, from whom all living people are descended patrilineally, perhaps even meeting and mating with him. Even if this were true, which is currently regarded as highly unlikely, this would only be a coincidence. Like mitochondrial "Eve", Y-chromosomal "Adam" probably lived in Africa. A recent study (March 2013) concluded however that "Eve" lived much later than "Adam" – some 140,000 years later.[7] (Earlier studies considered, conversely, that "Eve" lived earlier than "Adam".)[40] More recent studies indicate that mitochondrial Eve and Y-chromosomal Adam may indeed have lived around the same time.[41]
Not the most recent ancestor shared by all humans
Mitochondrial Eve is the most recent common matrilineal ancestor, not the most recent common ancestor. Since the mtDNA is inherited maternally and recombination is either rare or absent, it is relatively easy to track the ancestry of the lineages back to a MRCA; however, this MRCA is valid only when discussing mitochondrial DNA. An approximate sequence from newest to oldest can list various important points in the ancestry of modern human populations:
- The human MRCA. All humans alive today share a surprisingly recent common ancestor, perhaps even within the last 5,000 years, even for people born on different continents.[42]
- The identical ancestors point. Just a few thousand years before the most recent single ancestor shared by all living humans was the time at which all humans who were then alive either left no descendants alive today or were common ancestors of all humans alive today. In other words, "each present-day human has exactly the same set of genealogical ancestors" alive at the "identical ancestors point" in time. This is far more recent than when Mitochondrial Eve lived.[42]
- Mitochondrial Eve, the most recent female-line common ancestor of all living people.
- "Y-chromosomal Adam", the most recent male-line common ancestor of all living people.
Not the biblical Eve
Owing to its figurative reference to the first woman in the Biblical Book of Genesis, the Mitochondrial Eve theory initially met with enthusiastic endorsement from some young earth creationists, who viewed the theory as a validation of the biblical creation story. Some even went so far as to claim that the Mitochondrial Eve theory disproved evolution.[43][44][45] However, the theory does not suggest any relation between biblical Eve and Mitochondrial Eve because Mitochondrial Eve:
- is not a fixed individual (see above)
- had a mother
- was not the only woman of her time, and
- Y-chromosomal Adam is unlikely to have been her sexual partner, or indeed to have been contemporaneous to her.
Implications of dating and placement of Eve
There was initial resistance among some scientists to the Mitochondrial Eve hypothesis, in part because it challenged the widely accepted multiregional evolution hypothesis held by some leading paleoanthropologists, such as Milford Wolpoff. This prevailing theory held that the evolution of humanity from the beginning of the Pleistocene (2.5 million years ago) to the present day has been within a single, continuous human species, evolving worldwide to modern Homo sapiens. More resistance came from those who argued that there was too little time between Homo erectus and modern Homo sapiens to allow for another new species, and others who argued the regional evolution from archaic hominin forms into modern ones. Consequently, the finding of a recent maternal ancestor for all humans in Africa was very controversial.
Cann, Stoneking & Wilson (1987)'s placement of a relatively small population of humans in sub-Saharan Africa, lent appreciable support for the recent Out of Africa hypothesis. The current concept places between 1,500 and 16,000 effectively interbreeding individuals (census 4,500 to 48,000 individuals) within Tanzania and proximal regions. Later, Tishkoff et al. (2009) using data from many loci (not just mitochondrial DNA) extrapolated that the Angola-Namibia border region near the Atlantic Ocean is likely to be near the geographical point of origin of modern human genetic diversity. In its relatively southern origin proposals, this autosomal study was considered by the authors to be broadly consistent with previous mitochondrial DNA studies, including one by some of the same authors who associated the origins of mitochondrial haplogroups L0 and L1 with "click languages" in southern and eastern Africa.[46]
To some extent the studies have already revealed that the archaic Homo sapiens present in Northwest Africa (Jebel Irhoud) were not likely part of the contiguous modern human population. Better defined is the genetic separation among Neanderthals, Flores hobbit, Java man, and Peking man. In 1999 Krings et al. eliminated problems in molecular clocking postulated by Nei, 1992 when it was found that the mtDNA sequence for the same region was substantially different from the MRCA relative to any human sequence. Currently there are 6 fully sequenced Neanderthal mitogenomes, each falling within a genetic cluster less diverse than that for humans, and mitogenome analysis in humans has statistically markedly reduced the TMRCA range so that it no longer overlaps with Neandertal/human split times. Of all the non-African hominids, European archaics most closely resembled humans, indicating a wider genetic divide with other hominids.
Since the multiregional evolution hypothesis (MREH) revolved around a belief that regional modern human populations evolved in situ in various regions (Europe: Neandertals to Europeans, Asia: Homo erectus to East Asians, Australia: Sumatran erectines to indigenous Australians), these results demonstrated that a pure MREH hypothesis could not explain one important genetic marker.
In popular science
- Newsweek reported on Mitochondrial Eve based on the Cann et al. study in January 1988, under a heading of "Scientists Explore a Controversial Theory About Man's Origins". The edition sold a record number of copies.[47]
- The Seven Daughters of Eve presents the theory of human mitochondrial genetics to a general audience.[48]
- In River Out of Eden, Richard Dawkins discusses human ancestry in the context of a river of genes and shows that Mitochondrial Eve is one of the many common ancestors we can trace back to via different gene pathways.[49]
- The Discovery Channel produced a documentary entitled The Real Eve: 6 Billion People ... from ONE Woman (or Where We Came From in the United Kingdom), based on the book The Real Eve: Modern Man's Journey Out of Africa" by Stephen Oppenheimer.[50]
See also
|
Phylogenetic tree of human mitochondrial DNA (mtDNA) haplogroups | |||||||||||||||||||||||||||||||||||||
| Mitochondrial Eve (L) | |||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||
| CZ | D | E | G | Q | O | A | S | R | I | W | X | Y | |||||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | U | ||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||
| H | V | J | T | ||||||||||||||||||||||||||||||||||
References
- ↑ "the synonymous mutation rate of Kivisild et al. [...] estimates the coalescence time of the mtDNA tree overall at ~160,000 kya [...] We present a revised chronology using the complete mtDNA genome rate and an ML approach for the mtDNA tree in Figure 6, with full details of the age estimates and associated 95% confidence regions in Table S5. " Pedro Soares et al 2009, Correcting for Purifying Selection: An Improved Human Mitochondrial Molecular Clock. and its Supplemental Data. The American Journal of Human Genetics, Volume 84, Issue 6, 740–759, 4 June 2009. "we estimate the time to the most recent common ancestor (TMRCA) of the Y chromosome to be 120 to 156 thousand years and the mitochondrial genome TMRCA to be 99 to 148 thousand years. Our findings suggest that, contrary to previous claims, male lineages do not coalesce significantly more recently than female lineages." Poznik GD, Henn BM, Yee MC, Sliwerska E, Euskirchen GM, Lin AA, Snyder M, Quintana-Murci L, Kidd JM, Underhill PA, Bustamante CD (August 2013). "Sequencing Y chromosomes resolves discrepancy in time to common ancestor of males versus females". Science. 341 (6145): 562–565. doi:10.1126/science.1237619. PMID 23908239.
- ↑ Endicott, P; Ho, SY; Metspalu, M; Stringer, C (September 2009), "Evaluating the mitochondrial timescale of human evolution", Trends Ecol. Evol. (Amst.), 24 (9): 515–21, doi:10.1016/j.tree.2009.04.006, PMID 19682765
Soares P, Ermini L, Thomson N, et al. (June 2009), "Correcting for purifying selection: an improved human mitochondrial molecular clock", Am. J. Hum. Genet., 84 (6): 740–59, doi:10.1016/j.ajhg.2009.05.001, PMC 2694979
 , PMID 19500773.
University of Leeds – New 'molecular clock' aids dating of human migration history
, PMID 19500773.
University of Leeds – New 'molecular clock' aids dating of human migration history - 1 2 Soares, P; Ermini, L; Thomson, N; Mormina, M; Rito, T; Röhl, A; Salas, A; Oppenheimer, S; Macaulay, V (June 2009), "Correcting for purifying selection: an improved human mitochondrial molecular clock", American Journal of Human Genetics, 84 (6): 740–59, doi:10.1016/j.ajhg.2009.05.001, PMC 2694979, PMID 19500773
- 1 2 3 Poznik GD, Henn BM, Yee MC, Sliwerska E, Euskirchen GM, Lin AA, Snyder M, Quintana-Murci L, Kidd JM, Underhill PA, Bustamante CD (August 2013). "Sequencing Y chromosomes resolves discrepancy in time to common ancestor of males versus females". Science. 341 (6145): 562–565. doi:10.1126/science.1237619. PMID 23908239.
- 1 2 Dawkins, Richard (2004), The ancestor's tale: a pilgrimage to the dawn of evolution, Boston: Houghton Mifflin, ISBN 0-618-00583-8
- ↑ Mendez, Fernando; Krahn, Thomas; Schrack, Bonnie; Krahn, Astrid-Maria; Veeramah, Krishna; Woerner, August; Fomine, Forka Leypey Mathew; Bradman, Neil; Thomas, Mark (7 March 2013), "An African American paternal lineage adds an extremely ancient root to the human Y chromosome phylogenetic tree" (PDF), American Journal of Human Genetics, 92 (3): 454–9, doi:10.1016/j.ajhg.2013.02.002, PMC 3591855, PMID 23453668
- 1 2 Barras, Colin (6 March 2013). "The father of all men is 340,000 years old". New Scientist. Retrieved 13 March 2013.
- ↑ Francalacci P, Morelli L, Angius A, Berutti R, Reinier F, Atzeni R, Pilu R, Busonero F, Maschio A, Zara I, Sanna D, Useli A, Urru MF, Marcelli M, Cusano R, Oppo M, Zoledziewska M, Pitzalis M, Deidda F, Porcu E, Poddie F, Kang HM, Lyons R, Tarrier B, Gresham JB, Li B, Tofanelli S, Alonso S, Dei M, Lai S, Mulas A, Whalen MB, Uzzau S, Jones C, Schlessinger D, Abecasis GR, Sanna S, Sidore C, Cucca F (2013). "Low-pass DNA sequencing of 1200 Sardinians reconstructs European Y-chromosome phylogeny". Science. 341 (6145): 565–569. doi:10.1126/science.1237947. PMID 23908240.
- ↑ Cann RL (2013). "Genetics. Y weigh in again on modern humans". Science. 341 (6145): 465–467. doi:10.1126/science.1242899. PMID 23908212.
- ↑ "Jordan: 'Mitochondrial Eve'". weber.ucsd.edu. 2011. Retrieved 7 January 2012.
- ↑ Brown WM, George M Jr, Wilson AC; George; Wilson (1979), "Rapid evolution of animal mitochondrial DNA", Proc Natl Acad Sci U S A, 76 (4): 1967–1971, Bibcode:1979PNAS...76.1967B, doi:10.1073/pnas.76.4.1967, PMC 383514, PMID 109836
- ↑ Ferris SD, Brown WM, Wilson AC; Wilson; Brown (1981), "Evolutionary tree for apes and humans based on cleavage maps of mitochondrial DNA", Proc Natl Acad Sci U S A, 78 (4): 2432–2436, Bibcode:1981PNAS...78.2432F, doi:10.1073/pnas.78.4.2432, PMC 319360, PMID 6264476
- ↑ Brown WM (1980), "Polymorphism in mitochondrial DNA of humans as revealed by restriction endonuclease analysis", Proc Natl Acad Sci U S A, 77 (6): 3605–3609, doi:10.1073/pnas.77.6.3605, PMC 349666, PMID 6251473
- ↑ Cann RL, Brown WM, Wilson AC (1982), "Evolution of human mitochondrial DNA: a preliminary report", Prog Clin Biol Res, 103 (Pt A): 157–165, PMID 6298804
- ↑ Cann RL, Wilson AC (1983), "Length mutations in human mitochondrial DNA", Genetics, 104 (4): 699–711, PMC 1202135, PMID 6311667
- 1 2 Gitschier J (2010), "All about Mitochondrial Eve: An interview with Rebecca Cann", PLoS Genetics, 6 (3): e1000959, doi:10.1371/journal.pgen.1000959, PMC 2877732, PMID 20523888
- ↑ Denaro M, Blanc H, Johnson MJ, Chen KH, Wilmsen E, Cavalli-Sforza LL, Wallace DC (1981), "Ethnic variation in Hpa 1 endonuclease cleavage patterns of human mitochondrial DNA" (PDF), Proc Natl Acad Sci U S A, 78 (9): 5768–5772, Bibcode:1981PNAS...78.5768D, doi:10.1073/pnas.78.9.5768, PMC 348856, PMID 6272318
- ↑ Cann RL, Stoneking M, Wilson AC (1987), "Mitochondrial DNA and human evolution", Nature, 325 (6099): 31–36, Bibcode:1987Natur.325...31C, doi:10.1038/325031a0, PMID 3025745
- ↑ Vigilant L, Stoneking M, Harpending H, Hawkes K, Wilson AC (September 1991), "African populations and the evolution of human mitochondrial DNA", Science, 253 (5027): 1503–1507, Bibcode:1991Sci...253.1503V, doi:10.1126/science.1840702, PMID 1840702
- 1 2 Lewin R (1987), "The unmasking of mitochondrial Eve", Science, 238 (4823): 24–26, Bibcode:1987Sci...238...24L, doi:10.1126/science.3116666, PMID 3116666
- ↑ Wainscoat J (1987), "Out of the garden of Eden", Nature, 325 (6099): 13, Bibcode:1987Natur.325...13W, doi:10.1038/325013a0, PMID 3796736
- ↑ Wilkins A (27 January 2012). "The scientists behind Mitochondrial Eve tell us about the "lucky mother" who changed human evolution forever". On the Human. Gawker Media 2013. Retrieved 2013-05-03.
- ↑ Cann RL (1997), "Chapter 4: Mothers, Labels, and Misogyny", in Hager LD, Women in Human Evolution, London, UK: Routledge, pp. 75–89, ISBN 9780415108331
- ↑ Tierney J (1992). "The Search for Adam and Eve". Newsweek. Carter G. Woodson Institute for Afro-American and African Studies. Retrieved 2013-05-13.
- ↑ Lemonick MD (26 January 1987). "Everyone's Genealogical Mother". Time. Time Inc. Retrieved 2013-05-13.
- ↑ Darlu P, Tassy P (1987). "Disputed African origin of human populations". Nature. 329 (6135): 111–112. Bibcode:1987Natur.329..111D. doi:10.1038/329111b0. PMID 3114640.
- ↑ Maddison DR (1991). "African Origin of human mitochondrial DNA reexamined". Systematic Zoology. 40 (3): 355–363. doi:10.2307/2992327.
- ↑ Nei M (1992). "Age of the common ancestor of human mitochondrial DNA". Mol Biol Evol. 9 (6): 1176–1178. PMID 1435241.
- ↑ Ayala FJ (1995). "The myth of Eve: molecular biology and human origins". Science. 270 (5244): 1930–1936. Bibcode:1995Sci...270.1930A. doi:10.1126/science.270.5244.1930. PMID 8533083.
- ↑ Templeton A (2002). "Out of Africa again and again". Nature. 416 (6876): 45–51. doi:10.1038/416045a. PMID 11882887.
- ↑ Templeton A (1997). "Out of Africa? What do genes tell us?". Current Opinion in Genetics & Development. 7 (6): 841–847. doi:10.1016/S0959-437X(97)80049-4. PMID 9468796.
- ↑ Holsinger K (29 September 2012). "(Mathematics of the coalescent) An example: Mitochondrial Eve". Holsinger Lab. Retrieved 2013-05-16.
- ↑ Cyran KA, Kimmel M (2010). "Alternatives to the Wright-Fisher model: the robustness of mitochondrial Eve dating". Theor Popul Biol. 78 (3): 165–172. doi:10.1016/j.tpb.2010.06.001. PMID 20600209.
- ↑ Fu Q, Mittnik A, Johnson PL, Bos K, Lari M, Bollongino R, Sun C, Giemsch L, Schmitz R, Burger J, Ronchitelli AM, Martini F, Cremonesi RG, Svoboda J, Bauer P, Caramelli D, Castellano S, Reich D, Pääbo S, Krause J (March 21, 2013). "A revised timescale for human evolution based on ancient mitochondrial genomes". Current Biology. 23 (7): 553–559. doi:10.1016/j.cub.2013.02.044.
- ↑ Giles, Richard E; Blanc, H; Cann, H M; Wallace, D C (1980), "Maternal inheritance of human mitochondrial DNA", PNAS, 77 (11): 6715–6719, Bibcode:1980PNAS...77.6715G, doi:10.1073/pnas.77.11.6715, PMC 350359, PMID 6256757
- ↑ Birky, C. William (2008), "Uniparental inheritance of organelle genes", Current Biology, 18 (16): R692–R695, doi:10.1016/j.cub.2008.06.049, PMID 18727899
- ↑ Gibbons, A (January 1998), "Calibrating the Mitochondrial Clock", Science, 279 (5347): 28–29, Bibcode:1998Sci...279...28G, doi:10.1126/science.279.5347.28, PMID 9441404
- ↑ There are sites in mtDNA (such as: 16129, 16223, 16311, 16362) that evolve more rapidly, have been noted to change within intragenerational timeframes – Excoffier & Yang (1999).
- ↑ Takahata, N (January 1993), "Allelic genealogy and human evolution", Mol. Biol. Evol., 10 (1): 2–22, PMID 8450756""
- ↑ Cruciani, F; Trombetta, B; Massaia, A; Destro-Bisol, G; Sellitto, D; Scozzari, R (10 June 2011), "A Revised Root for the Human Y Chromosomal Phylogenetic Tree: The Origin of Patrilineal Diversity in Africa", The American Journal of Human Genetics, 88 (6): 814–818, doi:10.1016/j.ajhg.2011.05.002, PMC 3113241, PMID 21601174
- ↑ http://www.nature.com/news/genetic-adam-and-eve-did-not-live-too-far-apart-in-time-1.13478
- 1 2 Rohde, DL; Olson, S; Chang, JT (September 2004), "Modelling the recent common ancestry of all living humans", Nature, 431 (7008): 562–6, Bibcode:2004Natur.431..562R, doi:10.1038/nature02842, PMID 15457259
- ↑ Wieland C (2005), "Mitochondrial Eve and biblical Eve are looking good: criticism of young age is premature", Journal of Creation, 19 (1): 57–59
- ↑ Nelson CW (1 April 2003). "Genetics and Biblical Demographic Events". answersingenesis.org. Answers in Genesis. Retrieved 2013-05-16.
- ↑ Oakes J. "Can the human arguments about mitochondrial Eve and y-chromosome Adam be extended to the animal world to test the reality of the flood of Noah?". Evidence for Christianity. Answers in Genesis. Retrieved 2013-05-16.
- ↑ Tishkoff et al. (2007); also see Behar et al. (2008) for example.
- ↑ Oppenheimer, Stephen (2004), The Real Eve: Modern Man's Journey Out of Africa, New York: Carroll & Graf, ISBN 0-7867-1334-8
- ↑ Bryan Sykes (2002), The Seven Daughters of Eve: The Science That Reveals Our Genetic Ancestry, W. W. Norton & Company, ISBN 978-0-393-32314-6
- ↑ Richard Dawkins (1995), River out of eden: a Darwinian view of life, Basic Books, ISBN 978-0-465-06990-3, retrieved 5 December 2011
- ↑ Stephen Oppenheimer (2003), The Real Eve: Modern Man's Journey Out of Africa, New York: Basic Books, ISBN 0-7867-1192-2, retrieved 5 December 2011
Further reading
- Atkinson, QD; Gray, RD; Drummond, AJ (January 2009), "Bayesian coalescent inference of major human mitochondrial DNA haplogroup expansions in Africa", Proceedings of the Royal Society B: Biological Sciences, 276 (1655): 367–73, doi:10.1098/rspb.2008.0785, PMC 2674340, PMID 18826938
- Ayala, F (1995), "The myth of Eve:molecular biology and human origin", Science, 270 (5244): 1930–1936, Bibcode:1995Sci...270.1930A, doi:10.1126/science.270.5244.1930, PMID 8533083
- Balloux, F; Handley, LJ; Jombart, T; Liu, H; Manica, A (2009), "Climate shaped the worldwide distribution of human mitochondrial DNA sequence variation", Proc Biol Sci., 276 (1672): 3447–55, doi:10.1098/rspb.2009.0752, PMC 2817182, PMID 19586946
- Behar, D; Villems; Soodyall; Blue-Smith; Pereira; Metspalu; Scozzari; Makkan; Tzur (May 2008), "The dawn of human matrilineal diversity", American Journal of Human Genetics, 82 (5): 1130–40, doi:10.1016/j.ajhg.2008.04.002, PMC 2427203, PMID 18439549
- Brown, WM (June 1980), "Polymorphism in mitochondrial DNA of humans as revealed by restriction endonuclease analysis", Proc. Natl. Acad. Sci. U.S.A., 77 (6): 3605–9, Bibcode:1980PNAS...77.3605B, doi:10.1073/pnas.77.6.3605, PMC 349666, PMID 6251473
- Cann, RL; Stoneking, M; Wilson, AC (1987), "Mitochondrial DNA and human evolution", Nature, 325 (6099): 31–6, Bibcode:1987Natur.325...31C, doi:10.1038/325031a0, PMID 3025745
- Cox, MP (August 2008), "Accuracy of molecular dating with the rho statistic: deviations from coalescent expectations under a range of demographic models", Hum. Biol., 80 (4): 335–57, doi:10.3378/1534-6617-80.4.335, PMID 19317593
- Dawkins, Richard (2004), The ancestor's tale: a pilgrimage to the dawn of evolution, Boston: Houghton Mifflin, ISBN 0-618-00583-8
- Endicott, P; Ho, SY (April 2008), "A Bayesian evaluation of human mitochondrial substitution rates", Am. J. Hum. Genet., 82 (4): 895–902, doi:10.1016/j.ajhg.2008.01.019, PMC 2427281, PMID 18371929
- Endicott, P; Ho, SY; Metspalu, M; Stringer, C (September 2009), "Evaluating the mitochondrial timescale of human evolution", Trends Ecol. Evol. (Amst.), 24 (9): 515–21, doi:10.1016/j.tree.2009.04.006, PMID 19682765
- Excoffier, L; Yang, Z (October 1999), "Substitution rate variation among sites in mitochondrial hypervariable region I of humans and chimpanzees", Mol. Biol. Evol., 16 (10): 1357–68, doi:10.1093/oxfordjournals.molbev.a026046, PMID 10563016
- Felsenstein, J (April 1992), "Estimating effective population size from samples of sequences: inefficiency of pairwise and segregating sites as compared to phylogenetic estimates", Genet. Res., 59 (2): 139–47, doi:10.1017/S0016672300030354, PMID 1628818
- Ferris, SD; Brown, WM; Davidson, WS; Wilson, AC (October 1981), "Extensive polymorphism in the mitochondrial DNA of apes", Proc. Natl. Acad. Sci. U.S.A., 78 (10): 6319–23, Bibcode:1981PNAS...78.6319F, doi:10.1073/pnas.78.10.6319, PMC 349030, PMID 6273863
- Gibbons, Anne (January 1998), "Calibrating the mitochondrial clock", Science, 279 (5347): 28–9, Bibcode:1998Sci...279...28G, doi:10.1126/science.279.5347.28, PMID 9441404
- Gonder, MK; Mortensen, HM; Reed, FA; de Sousa, A; Tishkoff, SA (December 2007), "Whole-mtDNA genome sequence analysis of ancient African lineages", Mol. Biol. Evol, 24 (3): 757–68, doi:10.1093/molbev/msl209, PMID 17194802
- Hazelwood, L; Steele, J (2004), "Spatial dynamics of human dispersals: Constraints on modelling and archaeological validation", Journal of Archaeological Science, 31 (6): 669–679, doi:10.1016/j.jas.2003.11.009
- Ho, SY; Larson, G (February 2006), "Molecular clocks: when times are a-changin'", Trends Genet., 22 (2): 79–83, doi:10.1016/j.tig.2005.11.006, PMID 16356585
- Ingman, M; Kaessmann, H; Pääbo, S; Gyllensten, U (December 2000), "Mitochondrial genome variation and the origin of modern humans", Nature, 408 (6813): 708–13, doi:10.1038/35047064, PMID 11130070
- Kaessmann, H; Pääbo, S (January 2002), "The genetical history of humans and the great apes", J. Intern. Med., 251 (1): 1–18, doi:10.1046/j.1365-2796.2002.00907.x, PMID 11851860
- Loogväli, Eva-Liis; Kivisild, Toomas; Margus, Tõnu; Villems, Richard; O'Rourke, Dennis; et al. (2009), O'Rourke, Dennis, ed., "Explaining the Imperfection of the Molecular Clock of Hominid Mitochondria", PLoS ONE, 4 (12): e8260, Bibcode:2009PLoSO...4.8260L, doi:10.1371/journal.pone.0008260, PMC 2794369, PMID 20041137
- Kimura, Motoo; Ohta, Tomoko (2001), Theoretical Aspects of Population Genetics, Princeton University Press, p. 232, ISBN 0-691-08098-4
- Loewe, L; Scherer, S (November 1997), "Mitochondrial Eve: The Plot Thickens", Trends in Ecology & Evolution, 12 (11): 422–3, doi:10.1016/S0169-5347(97)01204-4
- Maca-Meyer, N; González, AM; Larruga, JM; Flores, C; Cabrera, VM (2001), "Major genomic mitochondrial lineages delineate early human expansions", BMC Genet., 2 (1): 13, doi:10.1186/1471-2156-2-13, PMC 55343, PMID 11553319
- Mishmar, D; Ruiz-Pesini, E; Golik, P; Macaulay, V; Clark, AG; Hosseini, S; Brandon, M; Easley, K; et al. (January 2003), "Natural selection shaped regional mtDNA variation in humans", Proc. Natl. Acad. Sci. U.S.A., 100 (1): 171–6, Bibcode:2002PNAS..100..171M, doi:10.1073/pnas.0136972100, PMC 140917, PMID 12509511
- Nei, M (November 1992), "Age of the common ancestor of human mitochondrial DNA", Mol. Biol. Evol., 9 (6): 1176–8, PMID 1435241
- Nielsen, R; Beaumont, MA (March 2009), "Statistical inferences in phylogeography", Mol. Ecol., 18 (6): 1034–47, doi:10.1111/j.1365-294X.2008.04059.x, PMID 19207258
- Oppenheimer, Stephen (2004), The Real Eve: Modern Man's Journey Out of Africa, New York: Carroll & Graf, ISBN 0-7867-1334-8
- Parsons, TJ; Muniec, DS; Sullivan, K; Alliston-Greiner, R; Wilson, MR; Berry, DL; Holland, KA; Weedn, VW; et al. (April 1997), "A high observed substitution rate in the human mitochondrial DNA control region", Nat. Genet., 15 (4): 363–8, doi:10.1038/ng0497-363, PMID 9090380
- Pritchard, JK; Seielstad, MT; Perez-Lezaun, A; Feldman, MW (1999), "Population growth of human Y chromosomes: a study of Y chromosome microsatellites.", Mol Biol Evol., 16 (12): 1791–98, doi:10.1093/oxfordjournals.molbev.a026091, PMID 10605120
- Reed, FA; Tishkoff, SA (2006), "Africa human diversity, origins and migrations", Current Opinion in Genetics & Development, 16: 598, doi:10.1016/j.gde.2006.10.008, PMID 17056248
- Rohde, DL; Olson, S; Chang, JT (September 2004), "Modelling the recent common ancestry of all living humans", Nature, 431 (7008): 562–6, Bibcode:2004Natur.431..562R, doi:10.1038/nature02842, PMID 15457259
- Santos, C; Sierra, B; Alvarez, L; Ramos, A; Fernández, E; Nogués, R; Aluja, MP (2008), "Frequency and pattern of heteroplasmy in the control region of human mitochondrial DNA", J Mol Evol., 67 (2): 191–200, doi:10.1007/s00239-008-9138-9, PMID 18618067
- Schaffner, SF (2004), "The X chromosome in population genetics", Nature Reviews Genetics, 5 (1): 43–51, doi:10.1038/nrg1247, PMID 14708015
- Soares, P; Ermini, L; Thomson, N; Mormina, M; Rito, T; Röhl, A; Salas, A; Oppenheimer, S; Macaulay, V (June 2009), "Correcting for purifying selection: an improved human mitochondrial molecular clock", American Journal of Human Genetics, 84 (6): 740–59, doi:10.1016/j.ajhg.2009.05.001, PMC 2694979, PMID 19500773
- Suissa, S; Wang, Z; Poole, J; Wittkopp, S; Feder, J; Shutt, TE; Wallace, DC; Shadel, GS; Mishmar, D (2009), Desalle, Rob, ed., "Ancient mtDNA genetic variants modulate mtDNA transcription and replication", PLoS Genet., 5 (5): e1000474, doi:10.1371/journal.pgen.1000474, PMC 2673036, PMID 19424428
- Sykes, Bryan (2001), The Seven Daughters of Eve, New York: Norton, ISBN 0-393-02018-5
- Sykes, Brian D. (2003), Adam's curse: a future without men, London: Bantam, ISBN 0-593-05005-3
- Takahata, N (January 1993), "Allelic genealogy and human evolution", Mol. Biol. Evol., 10 (1): 2–22, PMID 8450756
- Takahata, N; Lee, SH; Satta, Y (2001), "Testing multiregionality of modern human origins", Mol Biol Evol, 18 (2): 172–183, doi:10.1093/oxfordjournals.molbev.a003791, PMID 11158376
- Tamura, K,; Nei, M (May 1993), "Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees", Mol. Biol. Evol., 10 (3): 512–26, PMID 8336541
- Tang, H; Siegmund, DO; Shen, P; Oefner, PJ; Feldman, MW (May 2002), "Frequentist estimation of coalescence times from nucleotide sequence data using a tree-based partition", Genetics, 161 (1): 447–59, PMC 1462078, PMID 12019257*Vigilant, L; Pennington, R; Harpending, H; Kocher, TD; Wilson, AC (December 1989), "Mitochondrial DNA sequences in single hairs from a southern African population", Proc. Natl. Acad. Sci. U.S.A., 86 (23): 9350–4, Bibcode:1989PNAS...86.9350V, doi:10.1073/pnas.86.23.9350, PMC 298493, PMID 2594772
- Tishkoff, SA; -Gonder, MK; Henn, BM; Mortensen, H; Knight, A; Gignoux, C; Fernandopulle, N; Lema, G; Nyambo, TB (2007), "History of Click-Speaking Populations of Africa Inferred from mtDNA and Y Chromosome Genetic Variation" (PDF), Molecular Biology and Evolution, 24 (10): 2180–2195, doi:10.1093/molbev/msm155, PMID 17656633
- Tishkoff, S. A.; Reed, F. A.; Friedlaender, F. R.; Ehret, C.; Ranciaro, A.; Froment, A.; Hirbo, J. B.; Awomoyi, A. A.; Bodo, J.-M. (2009), "The Genetic Structure and History of Africans and African Americans", Science, 324 (5930): 1035–1044, Bibcode:2009Sci...324.1035T, doi:10.1126/science.1172257, PMC 2947357, PMID 19407144
- Vigilant, L; Stoneking, M; Harpending, H; Hawkes, K; Wilson, AC (September 1991), "African populations and the evolution of human mitochondrial DNA", Science, 253 (5027): 1503–7, Bibcode:1991Sci...253.1503V, doi:10.1126/science.1840702, PMID 1840702
- Watson E, Forster P, Richards M, Bandelt HJ (September 1997), "Mitochondrial footprints of human expansions in Africa", Am. J. Hum. Genet., 61 (3): 691–704, doi:10.1086/515503, PMC 1715955, PMID 9326335
- Wilson, AC; Cann, RL; Carr, SM; George M, UB; Gyllensten, KM; Helm-Bychowski, RG; Higuchi, SR; Palumbi, EM; Prager, Ellen M. (1985), "Mitochondrial DNA and two perspectives on evolutionary genetics", Biol J Linn Soc Lond., 26 (4): 375–400, doi:10.1111/j.1095-8312.1985.tb02048.x
- Wilder, JA; Mobasher, Z; Hammer, MF (2004), "Genetic evidence for unequal effective population sizes of human females and males", Mol Biol Evol., 21 (11): 2047–57, doi:10.1093/molbev/msh214, PMID 15317874
- White, TD; Asfaw, B; Beyene, Y; Haile-Selassie, Y; Lovejoy, CO; Suwa, G; WoldeGabriel, G (October 2009), "Ardipithecus ramidus and the paleobiology of early hominids", Science, 326 (5949): 75–86, Bibcode:2009Sci...326...75W, doi:10.1126/science.1175802, PMID 19810190
External links

| Wikimedia Commons has media related to Mitochondrial eve. |
- Krishna Kunchithapadam, "What, if anything, is a Mitochondrial Eve?" a simple explanation
- The Real Eve: Modern Man's Journey Out of Africa – by Stephen Oppenheimer – Discovery Channel, 2002
| Homininae | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Australopithecines |
| ||||||||||||||
| Humans and proto-humans |
| ||||||||||||||
| Related |
| ||||||||||||||