PDZ domain
|
Molecular structure of the PDZ domain included in the human GOPC (Golgi-associated PDZ and coiled-coil motif-containing protein) protein | |||||||||
| Identifiers | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Symbol | PDZ | ||||||||
| Pfam | PF00595 | ||||||||
| InterPro | IPR001478 | ||||||||
| SMART | PDZ | ||||||||
| PROSITE | PDOC50106 | ||||||||
| SCOP | 1lcy | ||||||||
| SUPERFAMILY | 1lcy | ||||||||
| CDD | cd00136 | ||||||||
| |||||||||

The PDZ domain is a common structural domain of 80-90 amino-acids found in the signaling proteins of bacteria, yeast, plants, viruses[1] and animals.[2] Proteins containing PDZ domains play a key role in anchoring receptor proteins in the membrane to cytoskeletal components. PDZ is an initialism combining the first letters of the first three proteins discovered to share the domain — post synaptic density protein (PSD95), Drosophila disc large tumor suppressor (Dlg1), and zonula occludens-1 protein (zo-1).[3] PDZ domains have previously been referred to as DHR (Dlg homologous region)[4] or GLGF (glycine-leucine-glycine-phenylalanine) domains.[5] Proteins with these domains help hold together and organize signaling complexes at cellular membranes. These domains play a key role in the formation and function of signal transduction complexes.[6] Protein domains, connected by intrinsically disordered flexible linker regions, induce long-range allostery via protein domain dynamics.[7] PDZ domains also play a highly significant role in the anchoring of cell surface receptors (such as Cftr and FZD7) to the actin cytoskeleton via mediators like NHERF and ezrin.[8]
In general PDZ domains bind to a short region of the C-terminus of other specific proteins. These short regions bind to the PDZ domain by beta sheet augmentation. This means that the beta sheet in the PDZ domain is extended by the addition of a further beta strand from the tail of the binding partner protein.[9]
Origins of discovery
PDZ is an acronym derived from the names of the first proteins in which the domain was observed. Post-synaptic density protein 95 (PSD-95) is a synaptic protein found only in the brain.[5] Drosophila disc large tumor suppressor (Dlg1) and zona occludens 1 (ZO-1) both play an important role at cell junctions and in cell signaling complexes.[10] Since the discovery of PDZ domains more than 20 years ago, researchers have successfully identified hundreds of PDZ domains. The first published use of the phrase “PDZ domain” was not in a paper, but a letter. In September 1995, Dr. Mary B. Kennedy of the California Institute of Technology wrote a letter of correction to Trends in Biomedical Sciences.[11] Earlier that year, another set of scientists had claimed to discover a new protein domain which they called a DHR domain.[12] Dr. Kennedy refuted that her lab had previously described exactly the same domain as a series of “GLGF repeats”.[5] She continued to explain that in order to “better reflect the origin and distribution of the domain,” the new title of the domain would be changed. Thus, the name “PDZ domain” was introduced to the world.
Structure
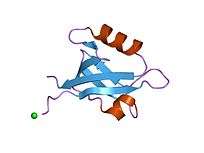
PDZ domain structure is not conserved across the various proteins that contain them, however there are similar trends seen in the PDZ domain. They usually have 6 β-strands and one short and one long α-helix. Apart from one fold that is conserved, the secondary structure is quite different across PDZ domain containing proteins.[6] This domain tends to have a globular confirmation of about 35 Å.[13]
When they are studied, PDZ domains are usually isolated into a monomer, however several studies have shown that some PDZ proteins form dimers. The function of PDZ protein dimers as compared to monomers is not yet known.[6]
The commonly accepted theory for the binding pocket of many PDZ domains is that it is made up of several hydrophobic amino acids. Most PDZ domains only have one binding site.[14]
Functions
PDZ domains have two main functions: Localizing cellular elements, and regulating cellular pathways.

The first discovered function of the PDZ domains was to anchor receptor proteins in the membrane to cytoskeletal components. PDZ domains also have regulatory functions on different signaling pathways.[15] Any protein may have one or several PDZ domains, which can be identical or unique (see figure to right). This variety allows these proteins to be very versatile in their interactions. Different PDZ domains in the same protein can have different roles, each binding a different part of the target protein or a different protein altogether.[16]
Localization
PDZ domains play a vital role in organizing and maintaining complex scaffolding formations, but this diversity in binding allows proteins with these domains to carry out a wide range of functions in this capacity.
PDZ domains are found in many different contexts and diverse proteins, but all assist in localization of cellular elements. PDZ domains are primarily involved in anchoring receptor proteins to the cytoskeleton. For cells to function properly it is important for the right components—proteins and other molecules— to be in the right place at the right time. Proteins with PDZ domains are able to bind to different components to ensure that the correct arrangement is met.[15] In the neuron, making sense of neurotransmitter activity requires specific receptors to be located in the lipid membrane at the synapse. PDZ domains are crucial to this receptor localization process.[17] Proteins with PDZ domains generally associate with both the C-terminus of the receptor and cytoskeletal elements in order to anchor the receptor to the cytoskeleton and keep it in place.[16][18] Without such an interaction, receptors would diffuse out of the synapse due to the fluid nature of the lipid membrane.
PDZ domains are also utilized to localize elements other than receptor proteins. In the human brain, nitric oxide often acts in the synapse to modify cGMP levels in response to NMDA receptor activation.[19] In order to ensure a favorable spatial arrangements, neuronal nitric oxide synthase (nNOS) is brought close to NMDA receptors via interactions with PDZ domains on PSD-95, which concurrently binds nNOS and NMDA receptors.[18] With nNOS located closely to NMDA receptors, it will be activated immediately after calcium ions begin entering the cell.
Regulation
PDZ domains are directly involved in the regulation of different cellular pathways. This mechanism of this regulation varies as PDZ domains are able to interact with a range of cellular components. This regulation is usually a result of the co-localization of multiple signaling molecules such as in the example with nNos and NMDA receptors.[18] Some examples of signaling pathway regulation executed by the PDZ domain include phosphatase enzyme activity, mecahnosensory signaling, and the sorting pathway of endocytosed receptor proteins.
The signaling pathway of the human protein tyrosine phosphatase non-receptor type 4 (PTPN4) is regulated by PDZ domains. This protein is involved in regulating cell death. Normally the PDZ domain of this enzyme is unbound. In this unbound state the enzyme is active and prevents cell signaling for apoptosis. Binding the PDZ domain of this phosphatase results in a loss of enzyme activity, which leads to apoptosis. The normal regulation of this enzyme prevents cells from prematurely going through apoptosis. When the regulation of the PTPN4 enzyme is lost, there is increased oncogenic activity as the cells are able to proliferate.[20]
PDZ domains also have a regulatory role in mechanosensory signaling in proprioceptors and vestibular and auditory hair cells. The protein Whirlin (WHRN) localizes in the post-synaptic neurons of hair cells that transform mechanical movement into action potentials that the body can interpret. WHRN proteins contains three PDZ domains. The domains located near the N-terminus bind to receptor proteins and other signaling components. When the one of these PDZ domains is inhibited, the signaling pathways of the neurons are disrupted, resulting in auditory, visual, and vestibular impairment. This regulation is thought to be based on the physical positioning WHRN and the selectivity of its PDZ domain.[21]
Regulation of receptor proteins occurs when the PDZ domain on the EBP50 protein binds to the C-terminus of the beta-2 adrenergic receptor (ß2-AR). EBP50 also associates with a complex that connects to actin, thus serving as a link between the cytoskeleton and ß2-AR. The ß2-AR receptor is eventually endocytosed, where it will either be consigned to a lysosome for degradation or recycled back to the cell membrane. Scientists have demonstrated that when the Ser-411 residue of the ß2-AR PDZ binding domain, which interacts directly with EBP50, is phosphorylated, the receptor is degraded. If Ser-411 is left unmodified, the receptor is recycled.[22] The role played by PDZ domains and their binding sites indicate a regulative relevance beyond simply receptor protein localization.
PDZ domains are being studied further to better understand the role they play in different signaling pathways. Research has increased as these domains have been linked to different diseases including cancer as discussed above.[23]
Regulation of PDZ Domain Activity
PDZ domain function can be both inhibited and activated by various mechanism. Two of the most prevalent include allosteric interactions and posttraslational modifications.[6]
Post-translational modifications
The most common post-traslational modification seen on PDZ domains is phosphorylation.[24] This modification is primarily an inhibitor of PDZ domain and ligand activity. In some examples, phosphorylation of amino acid side chains eliminates the ability of the PDZ domain to form hydrogen bonds, disrupting the normal binding patterns. The end result is a loss of PDZ domain function and further signaling.[25] Anther way phosphorylation can disrupt regualr PDZ domain funcition is by altering the charge ratio and further affecting binding and signaling.[26] In rare cases researchers have seen post-translational modifications activate PDZ domain activity[27] but these cases are few.

Another post-translational modification that can regulate PDZ domains is the formation of disulfide bridges. Many PDZ domains contain cysteines and are susceptible to disulfide bond formation in oxidizing conditions. This modification acts primarily as a inhibitor of PDZ domain function.[28]
Allosteric Interactions
Protein-protein interactions have been observed to alter the effectiveness of PDZ domains binding to ligands. These studies show that allosteric effects of certain proteins can affect the binding affinity for different substrates. Different PDZ domains can even have this allosteric effect on each other. This PDZ-PDZ interaction only acts as an inhibitor.[29] Other expiriments have shown that certain enzymes can enhance the binding of PDZ domains. Researchers found that the protein ezrin enhances the binding of the PDZ protein NHERF1.[8]
PDZ proteins
PDZ proteins are a family of proteins that contain the PDZ domain. This sequence of amino-acids is found in many thousands of known proteins. PDZ domain proteins are widespread in eukaryotes and eubacteria,[2] whereas there are very few examples of the protein in archaea. PDZ domains are often associated with other protein domains and these combinations allow them to carry out their specific functions. Three of the most well documented PDZ proteins are PSD-95, GRIP, and HOMER.
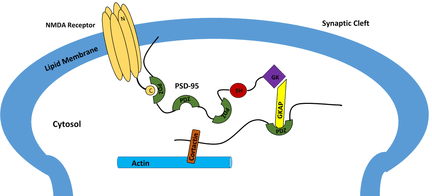
PSD-95 is a brain synaptic protein with three PDZ domains, each with unique properties and structures that allow PSD-95 to function in many ways. In general, the first two PDZ domains interact with receptors and the third interacts with cytoskeleton-related proteins. The main receptors associated with PSD-95 are NMDA receptors. The first two PDZ domains of PSD-95 bind to the C-terminus of NMDA receptors and anchor them in the membrane at the point of neurotransmitter release.[30] The first two PDZ domains can also interact in a similar fashion with Shaker-type K+ channels.[30] A PDZ interaction between PSD-95, nNOS and syntrophin is mediated by the second PDZ domain. The third and final PDZ domain links to cysteine-rich PDZ-binding protein (CRIPT), which allows PSD-95 to associate with the cytoskeleton.[30]
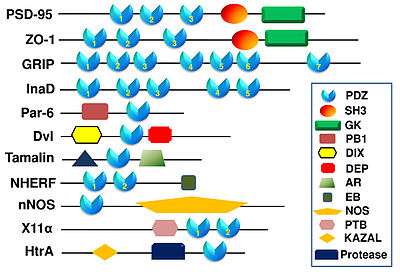
Glutamate receptor interacting protein (GRIP) is a post-synaptic protein with that interacts with AMPA receptors in a fashion analogous to PSD-95 interactions with NMDA receptors. When researchers noticed apparent structural homology between the C-termini of AMPA receptors and NMDA receptors, they attempted to determine if a similar PDZ interaction was occurring.[31] A yeast two-hybrid system helped them discover that out of GRIP’s seven PDZ domains, two (domains four and five) were essential for binding of GRIP to the AMPA subunit called GluR2.[16] This interaction is vital for proper localization of AMPA receptors, which play a large part in memory storage. Other researchers discovered that domains six and seven of GRIP are responsible for connecting GRIP to a family of receptor tyrosine kinases called ephrin receptors, which are important signaling proteins.[32] A clinical study concluded that Fraser syndrome, an autosomal recessive syndrome that can cause severe deformations, can be caused by a simple mutation in GRIP.[33]
HOMER differs significantly from many known PDZ proteins, including GRIP and PSD-95. Instead of mediating receptors near ion channels, as is the case with GRIP and PSD-95, HOMER is involved in metabotropic glutamate signaling.[34] Another unique aspect of HOMER is that it only contains a single PDZ domain, which mediates interactions between HOMER and type 5 metabotropic glutamate receptor (mGluR5).[17] The single GLGF repeat on HOMER binds amino acids on the C-terminus of mGluR5. HOMER expression is measured at high levels during embryologic stages in rats, suggesting an important developmental function.[17]
Human
There are roughly 260 PDZ domains in humans. Several proteins contain multiple PDZ domains, so the number of unique PDZ-containing proteins is closer to 180. Listed below are some of the better studied members of this family:
Below is a complete list:
AAG12; AHNAK; AHNAK2; AIP1; ALP; APBA1; APBA2; APBA3; ARHGAP21; ARHGAP23; ARHGEF11; ARHGEF12; CARD10; CARD11; CARD14; CASK; CLP-36; CNKSR2; CNKSR3; CRTAM; DFNB31; DLG1; DLG2; DLG3; DLG4; DLG5; DVL1; DVL1L1; DVL2; DVL3; ERBB2IP; FRMPD1; FRMPD2; FRMPD2L1; FRMPD3; FRMPD4; GIPC1; GIPC2; GIPC3; GOPC; GRASP; GRIP1; GRIP2; HTRA1; HTRA2; HTRA3; HTRA4; IL16; INADL; KIAA1849; LDB3; LIMK1; LIMK2; LIN7A; LIN7B; LIN7C; LMO7; LNX1; LNX2; LRRC7; MAGI1; MAGI2; MAGI3; MAGIX; MAST1; MAST2; MAST3; MAST4; MCSP; MLLT4; MPDZ; MPP1; MPP2; MPP3; MPP4; MPP5; MPP6; MPP7; MYO18A; NHERF1 ;NOS1; PARD3; PARD3B; PARD6A; PARD6B; PARD6G; PDLIM1; PDLIM2; PDLIM3; PDLIM4; PDLIM5; PDLIM7; PDZD11; PDZD2; PDZD3; PDZD4; PDZD5A; PDZD7; PDZD8; PDZK1; PDZRN3; PDZRN4; PICK1; PPP1R9A; PPP1R9B; PREX1; PRX; PSCDBP; PTPN13; PTPN3; PTPN4; RAPGEF2; RAPGEF6; RGS12; RGS3; RHPN1; RIL; RIMS1; RIMS2; SCN5A; SCRIB; SDCBP; SDCBP2; SHANK1; SHANK2; SHANK3; SHROOM2; SHROOM3; SHROOM4; SIPA1; SIPA1L1; SIPA1L2; SIPA1L3; SLC9A3R1; SLC9A3R2; SNTA1; SNTB1; SNTB2; SNTG1; SNTG2; SNX27; SPAL2; STXBP4; SYNJ2BP; SYNPO2; SYNPO2L; TAX1BP3; TIAM1; TIAM2; TJP1; TJP2; TJP3; TRPC4; TRPC5; USH1C; WHRN;
Virus
References
- ↑ Boxus M, Twizere JC, Legros S, Dewulf JF, Kettmann R, Willems L (2008). "The HTLV-1 Tax interactome". Retrovirology. 5: 76. doi:10.1186/1742-4690-5-76. PMC 2533353
 . PMID 18702816.
. PMID 18702816. - 1 2 Ponting CP (February 1997). "Evidence for PDZ domains in bacteria, yeast, and plants". Protein Sci. 6 (2): 464–468. doi:10.1002/pro.5560060225. PMC 2143646. PMID 9041651.
- ↑ Kennedy MB (September 1995). "Origin of PDZ(DHR,GLGF) domains". Trends Biochem. Sci. 20 (9): 350. doi:10.1016/S0968-0004(00)89074-X. PMID 7482701.
- ↑ Ponting CP, Phillips C (March 1995). "DHR domains in syntrophins, neuronal NO synthases and other intracellular proteins". Trends Biochem. Sci. 20 (3): 102–103. doi:10.1016/S0968-0004(00)88973-2. PMID 7535955.
- 1 2 3 Cho KO, Hunt CA, Kennedy MB (Nov 1992). "The rat brain postsynaptic density fraction contains a homolog of the Drosophila discs-large tumor suppressor protein". Neuron. 9 (5): 929–42. doi:10.1016/0896-6273(92)90245-9. PMID 1419001.
- 1 2 3 4 5 Lee HJ, Zheng JJ (2010). "PDZ domains and their binding partners: structure, specificity, and modification". Cell Commun. Signal. 8: 8. doi:10.1186/1478-811X-8-8. PMC 2891790. PMID 20509869.
- ↑ Bu Z, Callaway DJ (2011). "Proteins MOVE! Protein dynamics and long-range allostery in cell signaling". Advances in Protein Chemistry and Structural Biology. Advances in Protein Chemistry and Structural Biology. 83: 163–221. doi:10.1016/B978-0-12-381262-9.00005-7. ISBN 9780123812629. PMID 21570668.
- 1 2 Li, Jianquan; Callaway, David J.E.; Bu, Zimei (2009-09-11). "Ezrin induces long-range interdomain allostery in the scaffolding protein NHERF1". Journal of molecular biology. 392 (1): 166–180. doi:10.1016/j.jmb.2009.07.005. ISSN 0022-2836. PMC 2756645. PMID 19591839.
- ↑ Cowburn D (December 1997). "Peptide recognition by PTB and PDZ domains". Curr. Opin. Struct. Biol. 7 (6): 835–838. doi:10.1016/S0959-440X(97)80155-8. PMID 9434904.
- ↑ Liu, Jie; Li, Juan; Ren, Yu; Liu, Peijun (2014-01-01). "DLG5 in cell polarity maintenance and cancer development". International Journal of Biological Sciences. 10 (5): 543–549. doi:10.7150/ijbs.8888. ISSN 1449-2288. PMC 4046881. PMID 24910533.
- ↑ Kennedy, M. B. (1995-09-01). "Origin of PDZ (DHR, GLGF) domains". Trends in Biochemical Sciences. 20 (9): 350. doi:10.1016/s0968-0004(00)89074-x. ISSN 0968-0004. PMID 7482701.
- ↑ Ponting, Christopher P.; Phillips, Christopher (1995-03-01). "DHR domains in syntrophins, neuronal NO synthases and other intracellular proteins". Trends in Biochemical Sciences. 20 (3): 102–103. doi:10.1016/S0968-0004(00)88973-2.
- ↑ Erlendsson, Simon; Madsen, Kenneth Lindegaard (2015-10-16). "Membrane Binding and Modulation of the PDZ Domain of PICK1". Membranes. 5 (4): 597–615. doi:10.3390/membranes5040597. ISSN 2077-0375. PMC 4704001. PMID 26501328.
- ↑ Cabral, João H. Morais; Petosa, Carlo; Sutcliffe, Michael J.; Raza, Sami; Byron, Olwyn; Poy, Florence; Marfatia, Shirin M.; Chishti, Athar H.; Liddington, Robert C. (1996-08-15). "Crystal structure of a PDZ domain". Nature. 382 (6592): 649–652. doi:10.1038/382649a0.
- 1 2 Harris, B. Z.; Lim, W. A. (2001-09-01). "Mechanism and role of PDZ domains in signaling complex assembly". Journal of Cell Science. 114 (Pt 18): 3219–3231. ISSN 0021-9533. PMID 11591811.
- 1 2 3 Bristol, University of. "Bristol University | Centre for Synaptic Plasticity | AMPAR interactors". www.bristol.ac.uk. Retrieved 2015-12-03.
- 1 2 3 Brakeman, P. R.; Lanahan, A. A.; O'Brien, R.; Roche, K.; Barnes, C. A.; Huganir, R. L.; Worley, P. F. (1997-03-20). "Homer: a protein that selectively binds metabotropic glutamate receptors". Nature. 386 (6622): 284–288. doi:10.1038/386284a0. ISSN 0028-0836. PMID 9069287.
- 1 2 3 Doyle, Declan A.; Lee, Alice; Lewis, John; Kim, Eunjoon; Sheng, Morgan; MacKinnon, Roderick (1996-06-28). "Crystal Structures of a Complexed and Peptide-Free Membrane Protein–Binding Domain: Molecular Basis of Peptide Recognition by PDZ". Cell. 85 (7): 1067–1076. doi:10.1016/S0092-8674(00)81307-0.
- ↑ Hopper, Rachel; Lancaster, Barrie; Garthwaite, John (2004-04-01). "On the regulation of NMDA receptors by nitric oxide". European Journal of Neuroscience. 19 (7): 1675–1682. doi:10.1111/j.1460-9568.2004.03306.x. ISSN 1460-9568.
- ↑ Maisonneuve, Pierre; Caillet-Saguy, Célia; Raynal, Bertrand; Gilquin, Bernard; Chaffotte, Alain; Pérez, Javier; Zinn-Justin, Sophie; Delepierre, Muriel; Buc, Henri (2014-11-01). "Regulation of the catalytic activity of the human phosphatase PTPN4 by its PDZ domain". FEBS Journal. 281 (21): 4852–4865. doi:10.1111/febs.13024. ISSN 1742-4658.
- ↑ Nooij, Joriene C. de; Simon, Christian M.; Simon, Anna; Doobar, Staceyann; Steel, Karen P.; Banks, Robert W.; Mentis, George Z.; Bewick, Guy S.; Jessell, Thomas M. (2015-02-18). "The PDZ-Domain Protein Whirlin Facilitates Mechanosensory Signaling in Mammalian Proprioceptors". Journal of Neuroscience. 35 (7): 3073–3084. doi:10.1523/JNEUROSCI.3699-14.2015. ISSN 0270-6474. PMC 4331628. PMID 25698744.
- ↑ Cao, Tracy T.; Deacon, Heather W.; Reczek, David; Bretscher, Anthony; von Zastrow, Mark (1999-09-16). "A kinase-regulated PDZ-domain interaction controls endocytic sorting of the β2-adrenergic receptor". Nature. 401 (6750): 286–290. doi:10.1038/45816. ISSN 0028-0836.
- ↑ Wang, Nick X.; Lee, Ho-Jin; Zheng, Jie (2016-11-29). "Therapeutic use of PDZ protein-protein interaction antagonism". Drug news & perspectives. 21 (3): 137–141. ISSN 0214-0934. PMC 4055467. PMID 18560611.
- ↑ Chung, Hee Jung; Huang, Yan Hua; Lau, Lit-Fui; Huganir, Richard L. (2004-11-10). "Regulation of the NMDA receptor complex and trafficking by activity-dependent phosphorylation of the NR2B subunit PDZ ligand". The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 24 (45): 10248–10259. doi:10.1523/JNEUROSCI.0546-04.2004. ISSN 1529-2401. PMID 15537897.
- ↑ Jeleń, Filip; Oleksy, Arkadiusz; Smietana, Katarzyna; Otlewski, Jacek (2003-01-01). "PDZ domains - common players in the cell signaling". Acta Biochimica Polonica. 50 (4): 985–1017. ISSN 0001-527X. PMID 14739991.
- ↑ Chen, Jia; Pan, Lifeng; Wei, Zhiyi; Zhao, Yanxiang; Zhang, Mingjie (2008-08-06). "Domain-swapped dimerization of ZO-1 PDZ2 generates specific and regulatory connexin43-binding sites". The EMBO journal. 27 (15): 2113–2123. doi:10.1038/emboj.2008.138. ISSN 1460-2075. PMC 2516886. PMID 18636092.
- ↑ Chen, Bo-Shiun; Braud, Stephanie; Badger, John D.; Isaac, John T. R.; Roche, Katherine W. (2006-06-16). "Regulation of NR1/NR2C N-methyl-D-aspartate (NMDA) receptors by phosphorylation". The Journal of Biological Chemistry. 281 (24): 16583–16590. doi:10.1074/jbc.M513029200. ISSN 0021-9258. PMID 16606616.
- ↑ Mishra, Prashant; Socolich, Michael; Wall, Mark A.; Graves, Jennifer; Wang, ZiFen; Ranganathan, Rama (2007-10-05). "Dynamic scaffolding in a G protein-coupled signaling system". Cell. 131 (1): 80–92. doi:10.1016/j.cell.2007.07.037. ISSN 0092-8674. PMID 17923089.
- ↑ van den Berk, Lieke C. J.; Landi, Elena; Walma, Tine; Vuister, Geerten W.; Dente, Luciana; Hendriks, Wiljan J. A. J. (2007-11-27). "An allosteric intramolecular PDZ-PDZ interaction modulates PTP-BL PDZ2 binding specificity". Biochemistry. 46 (47): 13629–13637. doi:10.1021/bi700954e. ISSN 0006-2960. PMID 17979300.
- 1 2 3 Niethammer, M.; Valtschanoff, J. G.; Kapoor, T. M.; Allison, D. W.; Weinberg, R. J.; Craig, A. M.; Sheng, M. (1998-04-01). "CRIPT, a novel postsynaptic protein that binds to the third PDZ domain of PSD-95/SAP90". Neuron. 20 (4): 693–707. doi:10.1016/s0896-6273(00)81009-0. ISSN 0896-6273. PMID 9581762.
- ↑ Dong, H.; O'Brien, R. J.; Fung, E. T.; Lanahan, A. A.; Worley, P. F.; Huganir, R. L. (1997-03-20). "GRIP: a synaptic PDZ domain-containing protein that interacts with AMPA receptors". Nature. 386 (6622): 279–284. doi:10.1038/386279a0. ISSN 0028-0836. PMID 9069286.
- ↑ Torres, Richard; Firestein, Bonnie L; Dong, Hualing; Staudinger, Jeff; Olson, Eric N; Huganir, Richard L; Bredt, David S; Gale, Nicholas W; Yancopoulos, George D (1998-12-01). "PDZ Proteins Bind, Cluster, and Synaptically Colocalize with Eph Receptors and Their Ephrin Ligands". Neuron. 21 (6): 1453–1463. doi:10.1016/S0896-6273(00)80663-7.
- ↑ Vogel, Maartje J.; van Zon, Patrick; Brueton, Louise; Gijzen, Marleen; van Tuil, Marc C.; Cox, Phillip; Schanze, Denny; Kariminejad, Ariana; Ghaderi-Sohi, Siavash (2012-05-01). "Mutations in GRIP1 cause Fraser syndrome". Journal of Medical Genetics. 49 (5): 303–306. doi:10.1136/jmedgenet-2011-100590. ISSN 1468-6244. PMID 22510445.
- ↑ Ranganathan R, Ross E (1997). "PDZ domain proteins: scaffolds for signaling complexes". Curr Biol. 7 (12): R770–R773. doi:10.1016/S0960-9822(06)00401-5. PMID 9382826.
- ↑ Jemth P, Gianni S (July 2007). "PDZ domains: folding and binding". Biochemistry. 46 (30): 8701–8708. doi:10.1021/bi7008618. PMID 17620015.
Further reading
- Ponting CP, Phillips C, Davies KE, Blake DJ (June 1997). "PDZ domains: targeting signalling molecules to sub-membranous sites". BioEssays. 19 (6): 469–479. doi:10.1002/bies.950190606. PMID 9204764.
- Doyle DA, Lee A, Lewis J, Kim E, Sheng M, MacKinnon R (June 1996). "Crystal structures of a complexed and peptide-free membrane protein-binding domain: molecular basis of peptide recognition by PDZ". Cell. 85 (7): 1067–1076. doi:10.1016/S0092-8674(00)81307-0. PMID 8674113.
External links
- Eukaryotic Linear Motif resource motif class LIG_PDZ_Class_1
- Eukaryotic Linear Motif resource motif class LIG_PDZ_Class_2
- Eukaryotic Linear Motif resource motif class LIG_PDZ_Class_3
- The PDZ Domain as a Complex Adaptive System A concise technical summary and a statement of principal findings and ramifications of the PDZ Domain as a Complex Adaptive System
- NCBI conserved domains entry