Inbreeding depression
Inbreeding depression is the reduced biological fitness in a given population as a result of inbreeding, or breeding of related individuals. Population biological fitness refers to its ability to survive and reproduce itself. Inbreeding depression is often the result of a population bottleneck. In general, the higher the genetic variation or gene pool within a breeding population, the less likely it is to suffer from inbreeding depression.
Inbreeding depression seems to be present in most groups of organisms, but varies across mating systems. Hermaphroditic species often exhibit lower degrees of inbreeding depression than outcrossing species, as repeated generations of selfing is thought to purge deleterious alleles from populations. For example, the outcrossing nematode (roundworm) Caenorhabditis remanei has been demonstrated to suffer severely from inbreeding depression, unlike its hermaphroditic relative C. elegans, which experiences outbreeding depression.[2]
Mechanisms
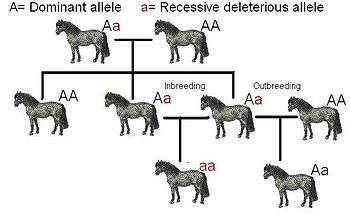
Inbreeding (i.e., breeding between closely related individuals) may on the one hand result in more recessive deleterious traits manifesting themselves, because the genomes of pair-mates are more similar: recessive traits can only occur in offspring if present in both parents' genomes, and the more genetically similar the parents are, the more often recessive traits appear in their offspring. Consequently, the more closely related the breeding pair is, the more homozygous deleterious genes the offspring may have, resulting in very unfit individuals. For alleles that confer an advantage in the heterozygous and/or homozygous-dominant state, the fitness of the homozygous-recessive state may even be zero (meaning sterile or unviable offspring).
An example of inbreeding depression is shown to the right. In this case, a is a recessive allele which has negative effects. In order for the a phenotype to become active, the gene must end up as aa because in the geneotype Aa, the A takes dominance over the a and the a does not have any effect. Due to their reduced phenotypic expression and their consequent reduced selection, recessive genes are, more often than not, detrimental phenotypes by causing the organism to be less fit to its natural environment.
Another mechanism responsible for inbreeding depression is the fitness advantage of heterozygous, which is known as overdominance. This can lead to reduced fitness of a population with many homozygous genotypes, even if they are not deleterious. Here, even the dominant alleles result in reduced fitness if present homozygously (see also hybrid vigour).
Currently, it is not known which of the two mechanisms is more prevalent in nature. For practical applications, e.g. in livestock breeding, the former is probably more significant – it may yield completely unviable offspring (meaning outright failure of a pedigree), while the latter can only result in relatively reduced fitness.
Inbreeding depression and natural selection
Natural selection cannot effectively remove all deleterious recessive genes from a population for several reasons. First, deleterious genes arise constantly through mutation within a population. Second, in a population where inbreeding occurs frequently, most offspring will have some deleterious traits, so few will be more fit for survival than the others. It should be noted that different deleterious traits are extremely unlikely to equally affect reproduction – an especially disadvantageous recessive trait expressed in a homozygous recessive individual is likely to eliminate itself, naturally limiting the expression of its phenotype. Third, recessive deleterious alleles will be "masked" by heterozygosity, and so in a dominant-recessive trait, heterozygotes will not be selected against.
When recessive deleterious alleles occur in the heterozygous state, where their potentially deleterious expression is masked by the corresponding wild-type allele, this masking phenomenon is referred to as complementation (see Complementation (genetics)).
In general, sexual reproduction in eukaryotes has two fundamental aspects: recombination during meiosis, and outcrossing. It has been proposed that these two aspects have two natural selective advantages respectively. A proposed adaptive advantage of meiosis is that it facilitates recombinational repair of DNA damages that are otherwise difficult to repair (see DNA repair as the adaptive advantage of meiosis). A proposed adaptive advantage of outcrossing is complementation, which is the masking of deleterious recessive alleles [3][4] (see hybrid vigor or heterosis). The selective advantage of complementation may largely account for the general avoidance of inbreeding (see Kin recognition).
Managing inbreeding depression
Introducing alleles from a different population can reverse inbreeding depression. Different populations of the same species have different deleterious traits, and therefore their crossbreeding will not result in homozygosity in most loci in the offspring. This is known as outbreeding enhancement, practiced by conservation managers and zoo captive breeders to prevent homozygosity.
However, intermixing two different populations may give rise to unfit polygenic traits in outbreeding depression, yielding offspring which lack the genetic adaptations to specific environmental conditions. These, then, will have a lowered fitness than pure-bred individuals e.g. of a particular subspecies that has adapted to its local environment.
In humans
The biological effects of inbreeding depression in humans are largely obscured by socioeconomic and cultural influences on reproductive behavior.[5] Studies in human populations have shown that age at marriage, duration of marriage, illiteracy, contraceptive use, and reproductive compensation are the major determinants of apparent fertility, even amongst populations with a high proportion of consanguinous unions.[6] However, several small - but significant - effects on increased mortality,[7] longer inter-birth intervals[7] and reduced overall productivity[5] have been noted in certain isolated populations.
Charles Darwin was one of the first scientists to demonstrate the effects of inbreeding depression, through numerous experiments on plants. Charles's wife, Emma, was his first cousin, and he was concerned about the impact of inbreeding on his ten children, three of whom died at age ten or younger; three others had childless long-term marriages.[8][9][10]
Factors reducing inbreeding depression
Whilst inbreeding depression has been found to occur in almost all sufficiently studied species, some taxa, most notably the angiosperms, appear to suffer lower fitness costs than others in inbred populations.[11] Three mechanisms appear to be responsible for this: purging, differences in ploidy and selection for heterozygosity.[11] It must be cautioned that some studies failing to show an absence of inbreeding depression in certain species can arise from small sample sizes or where the supposedly outbred control group is already suffering inbreeding depression, which frequently occurs in populations that have undergone a recent bottleneck, such as those of the naked mole rat.[11][12]
Purging selection
Purging selection occurs where the phenotypes of deleterious recessive alleles are exposed through inbreeding, and thus can be selected against. This can lead to such detrimental mutations being removed from the population, and has been demonstrated to occur rapidly where the recessive alleles have a lethal effect.[11] The efficiency of purging will depend on the relationship between the magnitude of the deleterious effect that is unmasked in the homozygotes and the importance of genetic drift, so that purging is weaker for non-lethal than for recessive lethal alleles.[13] For very small populations, drift has a strong influence, which can cause the fixation of sublethal alleles under weak selection.[11] The fixation of a single allele for a specific gene can also reduce fitness where heterozygote advantage was previously present (i.e., where heterozygous individuals have higher fitness than homozygotes of either allele), although this phenomenon seems to make a usually small contribution to inbreeding depression. Although naturally occurying purging can be important for population survival, deliberately attempting to purge deleterious mutations from a population is not generally recommended as a technique to improve the fitness of captive bred animals.[14][15][16]
Polyploidy
It has been noted that many angiosperms (flowering plants) can self-fertilise for several generations and suffer little from inbreeding depression. This is very useful for species which disperse widely and can therefore find themselves growing in a novel environment with no conspecifics present.[11] Polyploidy (having more than two paired sets of each chromosome), which is prevalent in angiosperms, ferns and a select few animal taxa, accounts for this. By having several copies of a chromosome, as opposed to two, homozygosity is less likely to occur in inbred offspring. This means that recessive deleterious alleles are not expressed as frequently as with many copies of a chromosome; it is more likely that at least one will contain a functional allele.[11]
Selection for heterozygosity
Inbreeding depression has also been found to occur more gradually than predicted in some wild populations, such as in the highly inbred population of Scandinavian wolves. This appears to be due to a selection pressure for more heterozygous individuals, which generally are in better condition and so are more likely to become one of the few animals to breed and produce offspring.[17]
See also
- Inbreeding
- Outbreeding depression
- Hybrid vigour
- Genetic purging
- Minimum viable population
- Population genetics
- Backward evolution
- Vadoma
- Insular dwarfism
- Island gigantism
References
- ↑ Begon, Michael, Colin R. Townsend, and John L. Harper. Ecology: from individuals to ecosystems. 4th ed. Malden, MA: Blackwell Pub., 2006. Print.
- ↑ Dolgin, Elie S.; Charlesworth, Brian; Baird, Scott; Cutter, Asher D.; et al. (2007). "Inbreeding and Outbreeding Depression in Caenorhabditis Nematodes". Evolution. 61 (6): 1339–1352. doi:10.1111/j.1558-5646.2007.00118.x. PMID 17542844.
- ↑ Bernstein, H; Byerly, HC; Hopf, FA; Michod, RE (1985). "Genetic damage, mutation, and the evolution of sex". Science. 229 (4719): 1277–1281. doi:10.1126/science.3898363. PMID 3898363.
- ↑ Michod, R.E. Eros and Evolution: A Natural Philosophy of Sex. (1996) Perseus Books ISBN 0201442329 ISBN 978-0201442328
- 1 2 http://www.nature.com/ejhg/journal/v17/n5/full/ejhg2008237a.html
- ↑ http://www.tandfonline.com/doi/abs/10.1080/03014460110075657
- 1 2 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1377721/
- ↑ Berra, Tim M.; Alvarez, Gonzalo; Ceballos, Francisco C.; et al. (2010). "Was the Darwin/Wedgwood Dynasty Adversely Affected by Consanguinity?". BioScience. 60 (5): 376–383. doi:10.1525/bio.2010.60.5.7.
- ↑ "Inbreeding May Have Caused Darwin Family Ills, Study Suggests". Science Daily.
- ↑ Clark, R.W. (1984) “The Survival of Charles Darwin” Random House [see pgs. 76 and 78]. ISBN 039452134X ISBN 978-0394521343
- 1 2 3 4 5 6 7 Frankham, Ballou, Briscoe (2002). Introduction to conservation genetics. Cambridge university press.
- ↑ Liberg, Paul; Firmin B. (2007). "Role of inbreeding depression and purging in captive breeding and restoration programmes". Molecular Ecology. 17 (1): 334–343. doi:10.1111/j.1365-294X.2007.03433.x. Retrieved 9 May 2014.
- ↑ García-Dorado, A (2012). "Understanding and predicting the fitness decline of shrunk populations: inbreeding, purging, mutation and standard selection". Genetics 190: 1461–1476. doi:10.1534/genetics.111.135541
- ↑ Garcia-Dorado, A (15 April 2015). "On the consequences of ignoring purging on geentic recommendations of MVP rules". Heredity. doi:10.1038/hdy.2015.28
- ↑ Leberg, P. L.; Firmin, B. D. (2008). "Role of inbreeding depression and purging in captive breeding and restoration programmes". Molecular Ecology. 17 (1): 334–343. doi:10.1111/j.1365-294X.2007.03433.x.
- ↑ Crnokrak, P; Barrett, SCH (2002). "Purging the genetic load: a review of the experimental evidence". Evolution. 56: 2347–2358. doi:10.1111/j.0014-3820.2002.tb00160.x.
- ↑ Bensch, Staffan; Andren, Hanson; Pederson, Sand; Sejberg, Wabakken; Akesson, Liberg (2006). "Selection for heterozygosity gives hope to wild wolves". PLOS ONE. 1: e72. doi:10.1371/journal.pone.0000072. PMC 1762340
 . PMID 17183704.
. PMID 17183704.