Genetic studies on Moroccans
Moroccan genetics encompasses the genetic history of the peoples of Morocco, and the genetic influence of this ancestry on world populations. It has been heavily influenced by geography.
In prehistoric times, the Sahara desert to the south and the Mediterranean Sea to the north were important geographical barriers. West Asia and Northeast Africa form a single land mass at the Suez. The Maghreb and Southwest Europe are separated by only 15 km (9 mi) at the Straits of Gibraltar, and a similar distance separates the Horn of Africa from the Arabian peninsula at the Bab el-Mandeb strait. At periods of low sea-levels, such as during a glacial maximum, islands that are currently submerged in the Mediterranean and possibly in between the Gibraltar straits would have been habitable. These may have encouraged bidirectional demic diffusion between Africa and Europe. During wetter phases of the Sahara, some populations in central Africa would have moved north into southern parts of North Africa. West Asian populations would have also been attracted to a wet Sahara, arriving either through the Suez, the Bab el-Mandeb or the Mediterranean.
As a result of these geographic influences, the genetic profile of the Moroccan population is a complex mosaic of autochthonous Maghrebi lineages, as well as Northeast African, European, West Asian and West African elements in variable degrees. Though Northwest Africa has experienced gene flow from the surrounding regions, it has also undergone long periods of genetic isolation in some parts. This has allowed distinctive genetic markers to evolve in some Maghrebi populations, especially in certain isolated Berber-speaking groups.
Prehistory and antiquity
The area of present-day Morocco has been inhabited since Paleolithic times, sometime between 90,000 and 190,000 BC.[1] During the Upper Paleolithic, the Maghreb was more fertile than it is today, resembling a savanna more than today's arid landscape.[2] 22,000 years ago, the Aterian was succeeded by the Iberomaurusian culture, which shared similarities with Iberian cultures. Skeletal similarities have been observed between the Iberomaurusian Mechta-Afalou burials and European Cro-Magnon remains. The Iberomaurusian industry in Morocco was succeeded by the Capsian culture.

.jpg)


North Africa and Morocco were slowly drawn into the wider emerging Mediterranean world by the Phoenicians, who established trading colonies and settlements in the early Classical period.[3] Mogador was a Phoenician colony as early as the early 6th century BC.[4]
Morocco later became part of a North African empire headquartered in Carthage. The earliest known independent Moroccan state was the Berber kingdom of Mauretania under king Bocchus I. This kingdom in northern Morocco, not to be confused with the present state of Mauritania, dates at least to 110 BC.[5]
The Roman Empire controlled this region from the 1st century BC, naming it Mauretania Tingitana. Christianity was introduced in the 2nd century AD and gained converts in the Roman towns, among slaves and some Berber farmers.
In the 5th century AD, as the Roman Empire declined, the region was invaded from the north first by the Vandals and then by the Visigoths. In the 6th century AD, northern Morocco was nominally part of the East Roman, or Byzantine Empire. Throughout this time, the Berber inhabitants in the high mountains of the interior of Morocco remained unsubdued.
Early Islamic era
In 670 AD, the first Islamic conquest of the North African coastal plain took place under Uqba ibn Nafi, a general serving under the Umayyads of Damascus. The Umayyad Muslims brought their language, their system of government, and Islam to Morocco. Many of the Berbers slowly converted to Islam, mostly after Arab rule had receded. The first independent Muslim state in the area of modern Morocco was the Kingdom of Nekor, an emirate in the Rif Mountains. It was founded by Salih I ibn Mansur in 710, as a client state to the Rashidun Caliphate. After the outbreak of the Great Berber Revolt in 739, the Berbers formed other independent states such as the Miknasa of Sijilmasa and the Barghawata.
According to medieval legend, Idris ibn Abdallah had fled to Morocco after the Abbasids' massacre of his tribe in Iraq. He convinced the Awraba Berber tribes to break their allegiance to the distant Abbasid caliphs in Baghdad and he founded the Idrisid Dynasty in 788. The Idrisids established Fes as their capital and Morocco became a centre of Muslim learning and a major regional power. The Idrissids were ousted in 927 by the Fatimid Caliphate and their Miknasa allies. After Miknasa broke off relations with the Fatimids in 932, they were removed from power by the Maghrawa of Sijilmasa in 980.
Berber dynasties


From the 11th century onwards, a series of powerful Berber[6][7][8] dynasties arose. Under the Almoravid dynasty [9] and the Almohad dynasty, Morocco dominated the Maghreb, much of present-day Spain and Portugal, and the western Mediterranean region. In the 13th and 14th centuries the Merinids held power in Morocco and strove to replicate the successes of the Almohads by military campaigns in Algeria and Spain. They were followed by the Wattasids. In the 15th century, the Reconquista ended Muslim rule in central and southern Spain and many Muslims and Jews fled to Morocco.[10] Portuguese efforts to control the Atlantic coast in the 15th century did not greatly affect the interior of Morocco. According to Elizabeth Allo Isichei, "In 1520, there was a famine in Morocco so terrible that for a long time other events were dated by it. It has been suggested that the population of Morocco fell from 5 to under 3 million between the early sixteenth and nineteenth centuries."[11]
Sharifian dynasties
In 1549, the region fell to successive Arab dynasties claiming descent from the Islamic prophet, Muhammad: first the Saadi dynasty who ruled from 1549 to 1659, and then the Alaouite dynasty, who remained in power since the 17th century.

Under the Saadi Dynasty, the country repulsed Ottoman incursions and a Portuguese invasion at the battle of Ksar el Kebir in 1578. The reign of Ahmad al-Mansur brought new wealth and prestige to the Sultanate, and a large expedition to West Africa inflicted a crushing defeat on the Songhay Empire in 1591. However, managing the territories across the Sahara proved too difficult. After the death of al-Mansur the country was divided among his sons.
In 1666 Morocco was reunited by the Alaouite Dynasty, who have been the ruling house of Morocco ever since. Morocco was facing aggression from Spain and the Ottoman Empire lies pressing westward. The Alaouites succeeded in stabilizing their position, and while the kingdom was smaller than previous ones in the region, it remained quite wealthy. Against the opposition of local tribes Ismail Ibn Sharif (1672–1727) began to create a unified state.[12] With his Jaysh d'Ahl al-Rif (the Riffian Army) he seized Tangier from the English in 1684 and drove the Spanish from Larache in 1689.
Morocco was the first nation to recognize the fledgling United States as an independent nation in 1777.[13][14][15][16] In the beginning of the American Revolution, American merchant ships in the Atlantic Ocean were subject to attack by the Barbary pirates. On 20 December 1777, Morocco's Sultan Mohammed III declared that American merchant ships would be under the protection of the sultanate and could thus enjoy safe passage. The Moroccan-American Treaty of Friendship, signed in 1786, stands as the U.S.'s oldest non-broken friendship treaty.[17][18]
Genetic evidence
The genetic proximity observed between Moroccans and Southern Europeans is due to the fact that both these groups shared a common ancestor either in the Upper Paleolithic, and in the Neolithic or alternatively during history with the invasion and the occupation during nearly seven centuries of the Iberian Peninsula by Moorish troops.[19] A genetic study published in January 2012 stated that the indigenous North-west African ancestry appears most closely related to populations outside of Africa but "divergence between Moroccan people and Near Eastern/Europeans likely precedes the Holocene (>12,000 ya) and The Paleolithic (>40.000BC)."[20]
Recent studies make clear no significant genetic differences exist between Arabic and non-Arabic speaking populations, The human leukocyte antigen HLA DNA data suggest that most Moroccans are of a Berber origin and that Arabs who invaded North Africa and Spain in the 7th century did not substantially contribute to the gene pool.[21][22] The Moorish refugees from Spain settled in the coast-towns.[23] According to a 2000 article in European Journal of Human Genetics, Moroccans from North-Western Africa were genetically closer to Iberians than to Sub-Saharan Africans of Bantu Ethnicity and Middle Easterners.[24]
The different loci studied revealed close similarity between the Berbers and other north African groups, mainly with Moroccan Arabic-speakers, which is in accord with the hypothesis that the current Moroccan population has a strong Berber background.[25]
Various population genetics studies along with historians such as Gabriel Camps and Charles-André Julien lend support to the idea that the bulk of the gene pool of modern Northwest Africans, irrespective of linguistic group, is derived from the Berber populations of the pre-Islamic period.[26]
According to the X-Chromosome SNP analyses, the authors reported a high genetic homogeneity between berbers and Arabs in NW Africa, so they suggested that the Arabisation of this area was a cultural phenomenon, which did not imply a replacement of the ancestry population. Our results give support the hypothesis of an early settlement of northwest Africa. The original berber population seem to have received a low genetic influx from the surrounding areas. Different hypothesis have been suggested to explain the genetic differentiation of the Moroccan population. An initial genetic drift could have caused differences in allele frequency distribution that have not been re-established due to a certain level of geographic isolation. The Strait of Gibraltar has been described by several authors as an important genetic barrier. Even a certain level of genetic exchange probably occurred between NW Africa and the South of the Iberian Peninsula, sharp frequency changes have been described in this area. Also the Sahara desert has been suggested as responsible of the genetic isolation of NW African populations from Sub-Saharan populations. There is no consensus about the impact of the Neolithic demic diffusion in the Mediterranean area. According to our results, a low impact of the Neolithic expansions and/or later migration events on NW African populations would have occurred. X-Chromosome SNP analyses
Moroccan Y-DNA chromosome
E-M215
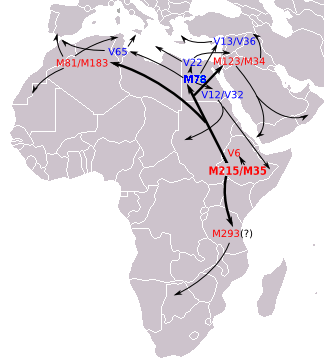
E1b1b (E-M215) is the most prevalent haplogroup in North Africa. E-M215 and its dominant subclade E-M35 is thought to have emerged in East-Africa about 22,400 years ago, and would have later dispersed into North Africa and from there into West Asia.[29][30] The E1b1b1 clade is presently found in various forms in Morocco. Total E1b1b1 (E-M35) frequencies reached at 93.8% in Moroccans.[31]
E-M215 has two ancient branches that contain all known modern E-M215 men, E-M35 and E-M281. Of these two, the only branch that has been confirmed in a native population outside of Ethiopia is E-M35, which in turn has four known branches, E-V68, E-Z827, E-V6 and E-V92. E-V68 and E-V257 have been found in highest numbers in North Africa and the Horn of Africa; but also in lower numbers in parts of the Middle East and Europe, and in isolated populations of Southern Africa.
E1b1b is quite common among Afro-Asiatic speakers. The linguistic group and carriers of its E-M35 lineage have a high probability to have arisen and dispersed together from the region of origin of this language family, amongst populations with an Afro-Asiatic speaking history.[32][33]
All major sub-branches of E-M35 are thought to have originated in the same general area as the parent clade: in North Africa, Horn of Africa, or nearby areas of the Near East. Some branches of E-M35 left Africa many thousands of years ago. For example, Battaglia et al. (2007) estimated that E-M78 (called E1b1b1a1 in that paper) has been in Europe longer than 10,000 years.
E-M81
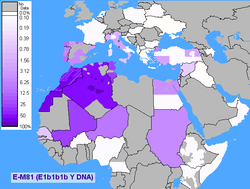
E1b1b1b1 (E-M81), formerly E1b1b1b, E3b1b, and E3b2, is the most common Y chromosome haplogroup in Morocco, dominated by its subclade E-M183.
It is thought to have originated in North Africa 14,200 years ago.[34] Its parent clade E1b1b (E-M215) is believed to have first appeared in Horn of Africa about 42,600 years ago.[35]
This haplogroup reaches a mean frequency of 85% in North Africa. It decreases in frequency from approximately 80% or more in some Moroccan Berber populations, including Saharawis, to approximately 10% to the east of this range in Egypt.[30][36]
Due to the clade's prevalence among these Berber groups and others such as Mozabites, Riffians, Chleuhs, Middle Atlas and Kabyle, it is sometimes referred to as a genetic Berber marker.
This phylogenetic tree of The Berber haplogroup subclades is based on the YCC 2008 tree and subsequent published research as summarized by ISOGG.[37][38][39]
- E1b1b1b (L19, V257)
- E1b1b1b1 (M81)
- E1b1b1b1a (M107) Underhill et al. (2000).
- E1b1b1b1b (M183) This clade is extremely dominant within E-M81. In fact, while Karafet et al. (2008) continues to describe this as a sub-clade of E-M81, and ISOGG defers to Karafet et al., all data seems to imply that it should actually be considered phylogenetically equivalent to M81
- E1b1b1b1b1 (M165) Underhill et al. (2000).
- E1b1b1b1b2 (L351) Found in two related participants in The E-M35 Phylogeny Project.
- E1b1b1b1 (M81)
Average North African Moroccan Berbers have frequencies of E3b3 in the +80%. Alvarez et al.(2009) study shows a frequency of E3b1b of 28/33 or 84.8% in Berbers from Marrakesh. With the rest of the frequencies being 1/33=3% E3a*, 1/33=3% E3b*, 1/33 or 3% E3b1a, and 1/33 or 3% E3b1c.[31]
E1b1b (M81) are Proto-Berber marker E1b1b1b1a1 (M107) A reduced Proto-Berber lineage in Mali.[40]
E-M78

The most basal and rare E-M78* paragroup has been found at lower frequencies in Moroccan Arabs. The sub-clade E-V65 is found in high levels in the Maghreb regions of far northern Africa. Cruciani et al. (2007) report levels of about 20% amongst Libyan Arab lineages, and about 30% amongst Moroccan Arabs. It appears to be less common amongst Berbers, but still present in levels of >10%. The authors suggest a North African origin for this lineage. In Europe, only a few individuals were found in Italy and Greece.
Capelli et al. (2009) studied the beta cluster in Europe. They found small amounts in Southern Italy, but also traces in Cantabria, Portugal and Galicia, with Cantabria having the highest level in Europe in their study, at 3.1% (5 out of 161 people).
Other frequencies of E1b1b1a1c (E-V22) is reported by Cruciani et al. (2007) include Moroccan Arabs (7.27%, 55 people) and Moroccan Jews (8%, 50 people).
Moroccan Y-DNA Haplogroups
| Population | n | A/B | E-M33 | E-V38 | E-M35* | E-M78 | E-M81 | E-M123 | G | J-P209 | R1 | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Morocco | 87 | — | — | 9.2 | — | 5.7 | 52.8 | — | — | 26.4 | — | Fadhlaoui-Zid et al. 2013[41] |
| Morocco | 221 | — | 1.8 | 4.5 | 4 | 6.8 | 65 | — | — | 9 | 4 | Fregel et al. 2009[42] |
| Morocco | 51 | 4 | 6 | — | 6 | 6 | 55 | — | — | 20 | 4 | Onofri et al. 2008[43] |
| Morocco | 176 | — | — | 6.3 | 5.1 | 6.3 | 63.6 | — | — | 13.6 | 2.8 | Bosch et al. 2001[44] |
| Arabs (Morocco) | 49 | — | — | — | — | 42.9 | 32.6 | — | — | 20.4 | — | Semino et al. 2004[45] |
| Arabs (Morocco) | 44 | — | — | 6.8 | 2.2 | 11.3 | 52.2 | — | — | 15.9 | 6.8 | Bosch et al. 2001[44] |
| Arabs (Morocco) | 54 | — | — | — | — | 38.9 | 31.5 | — | — | — | — | Cruciani et al. 2004[46] |
| Berbers (Morocco) | 64 | — | — | — | — | 10.9 | 68.7 | — | — | 6.3 | — | Semino et al. 2004[45] |
| Berbers (Marrakesh) | 29 | — | — | — | 3.4 | 6.9 | 72.4 | — | — | — | — | Cruciani et al. 2004[47] |
| Berbers (Middle Atlas) | 69 | — | — | — | — | 10.1 | 71 | — | 4.3 | 5.8 | — | Cruciani et al. 2004[47] |
| Berbers (Southern Morocco) | 40 | — | — | 2.5 | 7.5 | 12.5 | 65 | — | — | 10 | — | Bosch et al. 2001[44] |
| Berbers (North Central) | 63 | — | 3.1 | 9.5 | 7.9 | 1.5 | 65 | — | — | 11.1 | — | Bosch et al. 2001[44] |
| Berbers (Amizmiz) | 33 | 3 | — | — | 3 | 3 | 84.8 | 3 | — | — | — | Alvarez et al. 2009[48] |
| Berbers (Asni) | 54 | — | — | — | 1.9 | 3.7 | 79.6 | — | — | 1.9 | 1.9 | Dugoujon et al. (2005)[49] |
| Berbers (Bouhria) | 67 | — | — | — | — | 1.5 | 77.6 | — | 6 | 1.5 | 6 | Dugoujon et al. (2005)[49] |
| Berbers (Northern Morocco) | 43 | — | — | — | — | — | 79.1 | — | — | — | — | Ahmed Reguig et al. 2014[50] |
| Berbers (Southern Morocco) | 65 | — | — | — | — | — | 98.5 | — | — | — | — | Ahmed Reguig et al. 2014[50] |
| Berbers (Central Morocco) | 187 | — | — | — | — | — | 89.8 | — | — | — | — | Ahmed Reguig et al. 2014[50] |
| Moroccan Sahrawi | 89 | — | 8.9 | 11.2 | — | — | 59.5 | — | — | 20.2 | — | Fregel et al. 2009[42] |
| Moroccan Sahrawi | 29 | — | 3.4 | 3.4 | — | — | 75.8 | — | — | 17.2 | — | Bosch et al. 2001[44] |
| Moroccan Jews | 19 | — | — | — | 21.1 | — | — | — | 26.3 | 31.5 | 10.5 | Francalacci et al. 2008[51] |
J-P209
_Map.png)
Haplogroup J-P209 is believed to have arisen roughly 31,700 years ago in Southwest Asia (31,700±12,800 years ago according to Semino 2004). Haplogroup J-P209 is found in greatest concentration in Southwestern Arabian Peninsula. Outside of this region, haplogroup J-P209 has a presence in North Africa: Algeria (up to 35%) (Semino 2004), Tunisia (up to 31%),[52] Morocco (up to 20%) (Semino 2004), Egypt (up to 20%) (Luis 2004).
It also has a moderate presence in Southern Europe (especially in central and southern Italy, Malta, Greece, and Albania), Central Asia, and South Asia, particularly in the form of its subclade J-M172. Haplogroup J-P209 is also found in north East Africa, particularly in the form of its J-M267 subclade. The J-M410 subclade is found mostly in Greece, Anatolia, and southern Italy.
Other haplogroups
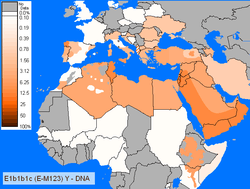
Concerning E-M123 without checking for the E-M34 SNP is found at small frequencies in Morocco A Low regional percentages for E-M123 was reported in Moroccan Berbers around 3%.
Eurasian haplogroups such as Haplogroup J and Haplogroup R1 have also been observed at very minimal frequencies. A thorough study by Cruciani et al. (2004) which analyzed populations from Morocco concludes that the North African pattern of Y-chromosomal variation (including both J1 and R1b haplogroups) is largely of Neolithic origin, which suggests that the Neolithic transition in this part of the world was accompanied by demic diffusion of Berber-speaking pastoralists from the Algerian Desert into Eastern Morocco, although later papers have suggested that this date could have been as long as ten thousand years ago, with the transition from the Oranian to the Capsian culture in North Africa.[53][54]
Haplogroups G and T are rarely found in Morocco, In 147 samples taken in Morocco, 1% were found to be G.[55]
In another study 1% of 312 samples in Morocco were G.[56]
Another study gathered samples only from hamlets in Morocco's Azgour Valley, where none of 33 samples were determined G.[31] These hamlets were selected because they were felt to be typically Berber in composition.
A study of 20 Moroccan Jews found 30% were G.[31] The tested men were then apparently living in Israel. Another study of Jewish men found 19.3% of 83 Jewish men from Morocco belonged to haplogroup G.[57] over G Moroccan samples are Likely Positive on the SNP G2a2b Haplogroup, it has been identified in neolithic human remains in Europe dating between 5000 and 3000BC. Furthermore, the majority of all the male skeletons from the European Neolithic period have so far yielded Y-DNA belonging to this haplogroup like the mummified remains of Ötzi the Iceman, The National Geographic Society places haplogroup G origins in the Middle East 30,000 years ago and presumes that people carrying the haplogroup took part in the spread of the Neolithic into Africa and then Europe [58] Two percent of Arab Moroccans and 0% to 8% of Berber Moroccans of Asni Oasis were likewise found to be G.[59]
Haplogroup T is found amongst central Berbers of Asni Oasis near the Algerian frontiers at 1,9% and observed in Moroccan Jews at 4%.
E1b1a is found at low frequencies in Morocco these lineages are found in some specific areas specially around the Great Desert Linked to the Slavery trade across the sahara like the presence of Haratins or Gnawa amongst Berbers of Asni Oasis located in North central Morocco near the Algerian frontiers, Sahrawis, Moroccan Arabs and in Southern Morocco.[60]
Haplogroup A1a is observed in southern and central Moroccan Berbers at 3%. related to the Homo-sapien Presence in North-west African Aterian and Mousterian Industries, one of The oldest Human branching event, is thought to have occurred about 140,000 years ago.[61]
The most basal and rare E1a* paragroup has been found at lower frequencies in samples obtained from Moroccan Berbers, and Sahrawis. dated around 45.000BC Linked to Back-Eurasian Migration from the Near East into North Africa along together with E1b1b during the Paleolithic times.[59]
Haplogroup distributions in Moroccan Populations

The major components of Y-DNA haplogroups present in Moroccan Berbers (E3b ; 94%) are shared with European and neighboring North African and Near Eastern populations. Minor share of haplogroups also include those related to North West Africans (E1a, A1a; 1%), Near Easterners (J, G, T; 2,4%), Sub Saharans Africans (E3a; 1,7%) and Europeans (R1b, I1; 2%) affinity.
Some of the major percentages identified were:
- E1b1b: 56% (Aboukhalid 2010) to 93.8% (Alvarez 2009) - Typical of people from the Mediterranean, North Africa, the Horn of Africa, and parts of the Arabian peninsula
- J-P209: 1.60% (El-Sibai 2009) to 20.4% (Semino 2004) - Typical of populations of the Arabian peninsula, the Levant and Caucasus, with a moderate distribution in Southeast Europe, North Africa, the Horn of Africa, Central Asia and South Asia.
- R1b: 0.8% to 6.8% (Bosch 2001) - Typical of Western Europeans, some West Asian peoples, the Sudanese Fulani and the Chadic-speaking peoples of Central Africa, and some Central Asian peoples (such as the Bashkirs, Turkmen and Uyghurs).[62]
- G: 0.4% - Typical of people from the Caucasus, and to a lesser extent the Middle East.
- A1a: 0.5% - Typical of Haratin-related groups in Northwest Africa.
- E1a: 0.5% - Rare haplogroup that has been found amongst Moroccan Berbers, Sahrawis, Southern Europeans, and some Chadic speakers in the Sahel.
- E1b1a: 1.7% - Typical of Niger-Congo-speaking populations.
- T: 0.4% - Typical of people from Northeast Africa, the Middle East and Central Asia..
- I: 0.4% - It can be found in the majority of present-day European populations, with peaks in Northern and South-Eastern Europe. Haplogroup I1 Y-chromosomes have also been found among some populations of the Near East, the Caucasus, Northeast Africa and Central Siberia.
Berber Genetic Identity of Moroccans

The prehistoric populations of Morocco, who were ancestral to Berbers, were related to the wider group of Paleo-Mediterranean peoples. The Afroasiatic family probably originated during the Mesolithic period, perhaps in the context of the Capsian culture.[63][64] DNA analysis has found commonalities between Berber Moroccan populations and those of the Sami people of Scandinavia showing a link dating from around 9,000 years ago.[65]
Around 5000 BC, the populations of North Africa were primarily descended from the makers of the Iberomaurusian and Capsian cultures, with a more recent intrusion associated with the Neolithic revolution.[66] The proto-Berber tribes evolved from these prehistoric communities during the Late Bronze to Early Iron Age.[67]
Genetic Prehistoric Expansions
DNA evidence suggests that during the Last Glacial Maximum, a period between 25,000 and 19,000 years ago, large ice sheets over a kilometer thick covered much of Northern Europe, making the region uninhabitable to humans. It is believed that human populations retreated south to warmer regions near the Mediterranean. Refuges during this period are believed to have been in Iberia, the Balkans and Italy. There was some gene flow from Morocco into Iberia.[68]
After the glacial maximum, when the European climate warmed up, the refuges are thought to have been the source from which Europe was repopulated. Berber lineages that had been introduced into the Iberian refuge would have then dispersed all over Europe with the Northward expansion of humans. This could explain the presence of genetic lineages in Eastern Europe and as far north as Russia, that appear to have prehistoric links to Northwest Africa, mainly Morocco (see mtDNA).[68] The expansion of human populations from Iberian refuges is also believed to have moved back to Morocco and Northwest Africa.[69]
Neolithic to the end of the prehistoric
The change from hunting and gathering to agriculture during the Neolithic Revolution was a watershed in world history. The societies that first made the change to agriculture are believed to have lived in North Africa and Middle East around 10,000 BCE. Agriculture was introduced into Europe by migrating farmers from the Middle East.[70] According to the demic diffusion model, these Middle Eastern farmers either replaced or interbred with the local hunter-gather populations that had been living in Europe since the "out of Africa" migration.[71]
It has been suggested that the first Middle Eastern farmers had North African influences mainly from The Capsian culture.[72] There have been suggestions that some genetic lineages found in the Middle East arrived there during this period.[73] The first Agricultural societies in the Middle East are generally thought to have emerged from the Natufian Culture, which existed in Palestine from 12,000 BCE-10,000 BCE. An important migration from North-West Africa occurred by the Ibero-Maurisians from Morocco across the Sinai appears to have occurred before the formation of the Natufian.
Genetic continuity in Morocco
In 2013, skeletons belonging to the makers of the Epipaleolithic Iberomaurusian culture, which were excavated at the prehistoric sites of Taforalt and Afalou, were analyzed for ancient DNA. All of the specimens belonged to maternal clades associated with either North Africa or the northern and southern Mediterranean littoral, indicating gene flow between these areas since the Epipaleolithic.[74] The ancient Taforalt individuals carried the mtDNA haplogroups U6, H, JT and V, which points to population continuity in the region dating from the Iberomaurusian period.[75]
Genetic Berber Heritage continuity of Moroccan Arabs
The cultural differentiation present in North Africa between Berber and Arab samples seems not to reflect genetic differences between both groups, as shown in the AMOVA analyses, and the MDS and PC analyses. If Arabs in Northern Africa were mostly descendants of Middle Eastern Arabs, the frequencies of haplogroups such as N, U1, U3, U7, and HV that are much more prevalent in the Middle East than elsewhere should be larger in North African Arabs than in Berbers. However, the opposite is observed : these haplogroups add up to 5% in North African Arabs but to 10% in Berbers.
The lack of differentiation between North African Arabs and Berbers has also been observed using other genetic markers such as classical markers (Bosch et al. 1997); autosomal STRs (Bosch et al. 2000), Alu insertion polymorphisms (Comas et al. 2000); and Y-chromosome lineages This pattern suggests that the Arabization of the area was mainly a cultural process, rather than a demographic replacement of the Berber populations that inhabited the region where the Arabic expansion took place.[76]
Moroccan Mitochondrial mtDNA
The Moroccan mitochondrial pool is essentially Berber in its structure, characterized by an "overall high frequency of Western Eurasian haplogroups" Represented by the Post-last glacial maximum expansion from Iberia to North Africa revealed by fine characterization of mtDNA HV haplogroup in Morocco is Estimated around 36% to 60%, a somehow lower frequency of sub-Saharan L lineages, and a significant (but differential) presence of North African haplogroups U6 and M1".[77] And according to Cherni et al. 2008 "the post-Last glacial maximum expansion originating in Iberia not only led to the resettlement of Europe but also of North Africa".[78]
Caucasoid mtDNA (maternal) sequences, were detected at frequencies of 96% in Moroccan Berbers, 82% in Algerian Berbers and 78% in non-Berber Moroccans, compared with only 4% in a Senegalese population. Rando 1998
Until recently, some papers suggested that the distribution of the main L haplogroups in Morocco was mainly due to trans-Saharan slave trade.[79] However, in September 2010, a thorough study about Berber mtDNA by Frigi et al. concluded that most of L haplogroups were much older and introduced by an ancient African gene flow around 20,000 years ago.[80]
in Otherways, Moroccan Northern and Southern Berbers have only 3% to 1% of SSA mtDNA, This north-south gradient in the sub-Saharan contribution to the gene pool is supported by Esteban et al.,[81] for the rest of mtDNA lineages mostly are Caucasian/West Eurasian, while Moroccan Arabs have more elevated SSA maternal admixture at around 21% to 36% Via L-mtDNA sequences, Highest frequencies of L-mtDNA is Reported to Moroccan Arabs of The Surrounding area of El jadida at 36% and this is largely ascribed to the slave trade.[82]
Frequencies (> 1%) of L-mtDNA
| Country | Ethnic Group | Number tested | Reference | L-mtDNA% |
| Morocco | Moroccan (Jews) | 149 | Behar et al. (2008) | 1.34% |
| Morocco | Moroccan Northern (berbers) | 124 | Esteban et al. (2004) | 1% |
| Morocco | Moroccan (Arabs) | 81 | Harich et al. (2010) | 36% |
| Morocco | Moroccan Arabs | 56 | Turchi et al. (2009) | 25.00% |
| Morocco | Moroccan Southern (Berbers) | 64 | Turchi et al. (2009) | 3.20% |
Genetic influence of Moroccans on Southern Europe
Trombetta et al. (2011) felt that V257 showed a parallel with its sibling clade E-V68 in the way that both clades show signs of having migrated from North West Africa Likely Morocco to southwestern Europe across the Mediterranean sea. They found 6 "E-V257*" individuals in their samples who were E-V257, from a Moroccan Marrakesh Berber, a Corsican, a Sardinian, a southern Spaniard and a Cantabrian.
Within E-M35, there are striking parallels between two haplogroups, E-V68 and E-V257. Both contain a lineage which has been frequently observed in North West Africa mainly Morocco (E-M78 and E-M81, respectively) and a group of undifferentiated chromosomes that are mostly found in southern Europe. An expansion of E-M35 carriers of E-V68* and E-V257* in the Northern Africa makes a maritime spread between Morocco and southern Europe a more plausible hypothesis.
A study from Semino (published 2004) showed that Y-chromosome haplotype E1b1b1b (E-M81), is specific to Moroccan populations and almost absent in Europe except Iberia (Spain and Portugal) and Sicily.
Y-chromosome analysis of the Iberian peninsula according to which haplogroup E1b1b1b surpasses frequencies of 10% in Southern Spain. The study points only to a very limited influence from northern Africa and the Middle East both in historic and prehistoric times.[83] The absence of microsatellite variation suggests a very recent arrival from Morocco consistent with historical exchanges across the Mediterranean during the period of Islamic expansion, namely of Berber populations. A study restricted to Portugal, concerning Y-chromosome lineages, revealed that "The mtDNA and Y data indicate that the Berber presence in that region dates prior to the Moorish expansion in 711 AD. ... Our data indicate that male Berbers, unlike sub-Saharan immigrants, constituted a long-lasting and continuous community in the country".[84]
Haplotype V(p49/TaqI), a characteristic Moroccan haplotype, may be also found in the Iberian peninsula, and a decreasing North-South cline of frequency clearly establishes a gene flow from Morocco towards Iberia which is also consistent with Moorish presence in the peninsula.[85] This North-South cline of frequency of halpotype V is to be observed throughout the Mediterranean region, ranging from frequencies of close to 30% in southern Portugal to around 10% in southern France. Similarly, the highest frequency in Italy is to be found in the southern island of Sicily (28%).[86][87]
A wide-ranging study (published 2007) using 6,501 unrelated Y-chromosome samples from 81 populations found that: "Considering both these E-M78 sub-haplogroups (E-V12, E-V22, E-V65) and the E-M81 haplogroup, the contribution of Moroccan lineages to the entire male gene pool of Iberia (barring Pasiegos), continental Italy and Sicily can be estimated as 5.6%, 3.6% and 6.6%, respectively."[87]
A study about Sicily by Gaetano et al. 2008 found that "The Hg E3b1b-M81, widely diffused in northwestern African Moroccan populations, is estimated to contribute to the Sicilian gene pool at a rate of 6%.".[88]
According to another recent study about Iberia by Adams et al. 2008 that analysed 1,140 unrelated Y-chromosome samples in Iberia, a much more important contribution of Moroccan lineages to the entire male gene pool of Iberia was found : "mean Moroccan admixture is 10.6%, with wide geographical variation, ranging from zero in Gascony to 21.7% in Northwest Castile".[55][89]
In Europe, E-M81 is found everywhere but mostly in the Iberian Peninsula, where unlike in the rest of Europe[Note 1] it is more common than E-M78, with an average frequency around 5%.[55] Its frequencies are higher in the western half of the peninsula with frequencies reaching 8% in Extremadura and South Portugal, 9% in Galicia, 10% in Western Andalusia and Northwest Castile and 9% to 17% in Cantabria.[55][90][91][92][93] The highest frequencies of this clade found so far in Europe were observed in the Pasiegos from Cantabria, ranging from 18% (8/45)[93] to 41% (23/56).[47] An average frequency of 8.28% (54/652) has also been reported in the Spanish Canary Islands with frequencies over 10% in the three largest islands of Tenerife (10.68%), Gran Canaria (11.54%) and Fuerteventura (13.33%).[94]
| Population | Moroccan admixture |
| Spain/Canary Island | 23.00% |
| Portugal | 15.40% |
| Spain/Galicia | 14.50% |
| Spain/Andalusia | 12.50% |
| Spain | 12.40% |
| France/Basque | 8.80% |
| Spain/Basque | 8.50% |
| France | 4.90% |
| Iberian region | %Moroccan male admixture |
| Castile, NorthWest | 21.7% |
| Minorca | 21.5% |
| Galicia | 20.8% |
| Extremadura | 19% |
| Andalucia, West | 16.7% |
| Portugal, South | 16.1% |
| Valencia | 12.8% |
| Portugal, North | 11.8% |
| Asturias | 10.5% |
| Castile, NorthEast | 9.3% |
| Majorca | 6.6% |
| Aragon | 4.8% |
| Ibiza | 3.8% |
| Andalucia, East | 2.4% |
| Catalonia | 2.3% |
| Castilla | 0.9% |
Genetic influences on Latin America
As a consequence of Spanish and Portuguese colonization of Latin America, E-M81 is also found throughout Latin America[95][96][97] and among Hispanic men in USA.[98]
Other regions
In other countries, Moroccan Berber haplogroups can be found in France, Sudan, Somalia, Jordan (4%),[91] Lebanon and amongst Sephardi Jews.
References
- ↑ Adams et al. 2008 shows an average frequency of 4.3% (49/1140) in the Iberian Peninsula with frequencies reaching 9% in Galicia, 10% in Western Andalusia and Northwest Castile. However this study also includes 153 individuals from Majorca, Minorca and Ibiza islands as well as 24 individuals from Gascony which are not in the Iberian Peninsula. Without these 177 individuals, real average for Iberian Peninsula is 4.9% (47/963), see table.
- ↑ Field Projects – Jebel Irhoud . Department of Human Evolution. Max Planck Institute for Evolutionary Anthropology
- ↑ Rubella, D. (1984). "Environmentalism and Pi Paleolithic economies in the Maghreb (c. 20,000 to 5000 B.P.)". In J.D. Clark & S.A. Brandt. From hunters to farmers the causes and consequences of food production in Africa. Berkeley: University of California Press. pp. 41–56. ISBN 0520045742.
- ↑ The Megalithic Portal; Megalith Map. "C. Michael Hogan, ''Mogador: Promontory Fort'', The Megalithic Portal, ed. Andy Burnham". Megalithic.co.uk. Retrieved 2010-06-02.
- ↑ Moscati, Sabatino (2001) The Phoenicians, Tauris, ISBN 1-85043-533-2
- ↑ "Bocchus I". Retrieved September 27, 2010.
- ↑ Ramirez-Faria, Carlos (2007-01-01). Concise Encyclopaedia of World History. ISBN 978-81-269-0775-5.
- ↑ "Almoravides". Universalis Encyclopedia.
- ↑ "Marīnid dynasty". Encyclopædia Britannica.
- ↑ "The Maghrib under the Almoravids and the Almohads". Britannica.com. Retrieved 2011-08-01.
- ↑ "Morocco – History". Britannica.com. Retrieved 2011-08-01.
- ↑ Allo Isichei, Elizabeth (1997). A history of African societies to 1870. Cambridge University Press. p. 264. ISBN 0-521-45599-5
- ↑ "Morocco (Page 8 of 9)". Microsoft Encarta Online Encyclopedia 2009. 2009-11-01.
- ↑ http://www.whitehouse.gov/the-press-office/2013/11/22/joint-statement-united-states-america-and-kingdom-morocco
- ↑ https://books.google.fr/books?id=5V77mdCXHJcC&pg=PA114
- ↑ http://www.defense.gov/news/newsarticle.aspx?id=41811
- ↑ Kozaryn, Linda D. "Cohen Renews U.S.-Morocco Ties". U.S. Department of Defense. Retrieved 2009-03-12.
- ↑ Roberts, Priscilla H. and Richard S. Roberts, Thomas Barclay (1728–1793): Consul in France, Diplomat in Barbary, Lehigh University Press, 2008, pp. 206–223 ISBN 093422398X.
- ↑ "Milestones of American Diplomacy, Interesting Historical Notes, and Department of State History". U.S. Department of State. Retrieved 2007-12-17.
- ↑ Coudray, C.; Olivieri, A.; Achilli, A.; Pala, M.; Melhaoui, M.; Cherkaoui, M.; El-Chennawi, F.; Kossmann, M.; Torroni, A.; Dugoujon, J. M. (2009). "The Complex and Diversified Mitochondrial Gene Pool of Berber Populations". Annals of Human Genetics. 73 (2): 196–214. doi:10.1111/j.1469-1809.2008.00493.x. PMID 19053990.
- ↑ Henn, B. M.; Botigué, L. R.; Gravel, S.; Wang, W.; Brisbin, A.; Byrnes, J. K.; Fadhlaoui-Zid, K.; Zalloua, P. A.; Moreno-Estrada, A. (2012). Schierup, Mikkel H, ed. "Genomic Ancestry of North Africans Supports Back-to-Africa Migrations". PLoS Genetics. 8 (1): e1002397. doi:10.1371/journal.pgen.1002397. PMC 3257290
 . PMID 22253600.
. PMID 22253600. - ↑ Ballais, Jean-Louis (2000) "Chapter 7: Conquests and land degradation in the eastern Maghreb" In Barker, Graeme and Gilbertson, David (2000) The Archaeology of Drylands: Living at the Margin Routledge, London, Volume 1, Part III – Sahara and Sahel, pp. 125–136, ISBN 978-0-415-23001-8
- ↑ Gomez-Casado, E.; Del Moral, P.; Martinez-Laso, J.; Garcia-Gomez, A.; Allende, L.; Silvera-Redondo, C.; Longas, J.; Gonzalez-Hevilla, M.; Kandil, M. (2000). "HLA genes in Arabic-speaking Moroccans: Close relatedness to Berbers and Iberians". Tissue Antigens. 55 (3): 239–249. doi:10.1034/j.1399-0039.2000.550307.x. PMID 10777099.
- ↑ Carr, Matthew (2009). Blood and faith: the purging of Muslim Spain. The New Press. p. 290. ISBN 1-59558-361-0.
- ↑ Bosch, E.; Calafell, F.; Pérez-Lezaun, A.; Clarimón, J.; Comas, D.; Mateu, E.; Martínez-Arias, R.; Morera, B.; Brakez, Z. (2000). "Genetic structure of north-west Africa revealed by STR analysis". European Journal of Human Genetics. 8 (5): 360–366. doi:10.1038/sj.ejhg.5200464. PMID 10854096.
- ↑ Harich, N.; Esteban, E.; Chafik, A.; López-Alomar, A.; Vona, G.; Moral, P. (2002). "Classical polymorphisms in Berbers from Moyen Atlas (Morocco): Genetics, geography, and historical evidence in the Mediterranean peoples". Annals of Human Biology. 29 (5): 473–487. doi:10.1080/03014460110104393. PMID 12396367.
- ↑ Arredi, B.; Poloni, E.; Paracchini, S.; Zerjal, T.; Fathallah, D.; Makrelouf, M.; Pascali, V.; Novelletto, A.; Tyler-Smith, C. (2004). "A Predominantly Neolithic Origin for Y-Chromosomal DNA Variation in North Africa". The American Journal of Human Genetics. 75 (2): 338–345. doi:10.1086/423147. PMC 1216069. PMID 15202071.
- ↑ Henn et al. (2008)
- ↑ Hassan et al. (2008)
- ↑ Cruciani, F.; La Fratta, R.; Santolamazza, P.; Sellitto, D.; Pascone, R.; Moral, P.; Watson, E.; Guida, V.; Colomb, E. B. (2004). "Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out of Africa". The American Journal of Human Genetics. 74 (5): 1014–1022. doi:10.1086/386294. PMC 1181964. PMID 15042509.
- 1 2 Arredi et al. (2004)
- 1 2 3 4 Alvarez, L.; Santos, C.; Montiel, R.; Caeiro, B.; Baali, A.; Dugoujon, J. M.; Aluja, M. P. (2009). "Y-chromosome variation in South Iberia: Insights into the North African contribution". American Journal of Human Biology. 21 (3): 407–409. doi:10.1002/ajhb.20888. PMID 19213004.
- ↑ Ehret et al. (2004)
- ↑ Keita & Boyce (2005)
- ↑ "E-M81 YTree". www.yfull.com. Retrieved 2016-07-10.
- ↑ "E-M215 YTree". www.yfull.com. Retrieved 2016-07-10.
- ↑ Alvarez et al. 2009
- ↑ ISOGG (2008)
- ↑ Karafet, T. M.; Mendez, F. L.; Meilerman, M. B.; Underhill, P. A.; Zegura, S. L.; Hammer, M. F. (2008). "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. 18 (5): 830–838. doi:10.1101/gr.7172008. PMC 2336805. PMID 18385274.
- ↑ Consortium, T. Y. C. (2002). "A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups". Genome Research. 12 (2): 339–348. doi:10.1101/gr.217602. PMC 155271. PMID 11827954.
- ↑ "The Genetic Atlas - E1b1b Meditid mutative history". thegeneticatlas.com. Retrieved 2015-10-01.
- ↑ Fadhlaoui-Zid et al. 2013, Genome-Wide and Paternal Diversity Reveal a Recent Origin of Human Populations in North Africa
- 1 2 Fregel et al. 2009, Demographic history of Canary Islands male gene-pool: replacement of native lineages by European.
- ↑ Onofri et al. 2008, Y-chromosome markers distribution in Northern Africa: High-resolution SNP and STR analysis in Tunisia and Morocco populations
- 1 2 3 4 5 Bosch et al. 2001, High-resolution analysis of humanY-chromosome variation shows a sharp discontinuity and limited gene flow between Northwestern Africa and the IberianPeninsula.
- 1 2 Semino, O.; Magri, C.; Benuzzi, G.; Lin, A. A.; Al-Zahery, N.; Battaglia, V.; MacCioni, L.; Triantaphyllidis, C.; Shen, P. (2004). "Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area". The American Journal of Human Genetics. 74 (5): 1023–34. doi:10.1086/386295. PMC 1181965. PMID 15069642.
- ↑ Cruciani et al. 2004, Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out Of Africa.
- 1 2 3 Cruciani, F.; La Fratta, R.; Santolamazza, P.; Sellitto, D.; Pascone, R.; Moral, P.; Watson, E.; Guida, V.; Colomb, E. B. (2004). "Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out of Africa". The American Journal of Human Genetics. 74 (5): 1014–1022. doi:10.1086/386294. PMC 1181964. PMID 15042509.
- ↑ Alvarez, Luis; Santos, Cristina; Montiel, Rafael; Caeiro, Blazquez; Baali, Abdellatif; Dugoujona, Jean-Michel; Dugoujon, Jean-Michel; Aluja, Maria Pilar (2009-06-01). "Y-chromosome variation in South Iberia: insights into the North African contribution". American Journal of Human Biology: The Official Journal of the Human Biology Council. 21 (3): 407–409. doi:10.1002/ajhb.20888. ISSN 1520-6300. PMID 19213004.
- 1 2 "The Berbers: Linguistic and genetic diversity" (PDF). ddl.ish-lyon.cnrs.fr. Retrieved 22 March 2015.
- 1 2 3 Ahmed, Reguig,; Nourdin, Harich,; Abdelhamid, Barakat,; Hassan, Rouba, (2014-01-01). "Phylogeography of E1b1b1b-M81 Haplogroup and Analysis of its Subclades in Morocco". 86 (2).
- ↑ Francalacci et al. 2008, History and geography of human Y-chromosome in Europe: a SNP perspective
- ↑ 31% is based on Combined Data
- Semino 2004: 30%
- Arredi 2004: 32%
- ↑ Semino, O.; Magri, C.; Benuzzi, G.; Lin, A. A.; Al-Zahery, N.; Battaglia, V.; MacCioni, L.; Triantaphyllidis, C.; Shen, P. (2004). "Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area". The American Journal of Human Genetics. 74 (5): 1023–34. doi:10.1086/386295. PMC 1181965. PMID 15069642.
- ↑ Myles, S.; Bouzekri, N.; Haverfield, E.; Cherkaoui, M.; Dugoujon, J. M.; Ward, R. (2005). "Genetic evidence in support of a shared Eurasian-North African dairying origin". Human Genetics. 117 (1): 34–42. doi:10.1007/s00439-005-1266-3. PMID 15806398.
- 1 2 3 4 Adams, S. M.; Bosch, E.; Balaresque, P. L.; Ballereau, S. P. J.; Lee, A. C.; Arroyo, E.; López-Parra, A. M.; Aler, M.; Grifo, M. S. G. (2008). "The Genetic Legacy of Religious Diversity and Intolerance: Paternal Lineages of Christians, Jews, and Muslims in the Iberian Peninsula". The American Journal of Human Genetics. 83 (6): 725–736. doi:10.1016/j.ajhg.2008.11.007. PMC 2668061. PMID 19061982.
- ↑ Zalloua, P. A.; Platt, D. E.; El Sibai, M.; Khalife, J.; Makhoul, N.; Haber, M.; Xue, Y.; Izaabel, H.; Bosch, E. (2008). "Identifying Genetic Traces of Historical Expansions: Phoenician Footprints in the Mediterranean". The American Journal of Human Genetics. 83 (5): 633–42. doi:10.1016/j.ajhg.2008.10.012. PMC 2668035. PMID 18976729.
- ↑ Behar, D. M.; Yunusbayev, B.; Metspalu, M.; Metspalu, E.; Rosset, S.; Parik, J. R.; Rootsi, S.; Chaubey, G.; Kutuev, I. (2010). "The genome-wide structure of the Jewish people". Nature. 466 (7303): 238–242. doi:10.1038/nature09103. PMID 20531471.
- ↑ Alonso, S.; Flores, C.; Cabrera, V.; Alonso, A.; Martín, P.; Albarrán, C.; Izagirre, N.; De La Rúa, C. N.; García, O. (2005). "The place of the Basques in the European Y-chromosome diversity landscape". European Journal of Human Genetics. 13 (12): 1293–1302. doi:10.1038/sj.ejhg.5201482. PMID 16094307.
- 1 2 Cruciani, F.; Santolamazza, P.; Shen, P.; MacAulay, V.; Moral, P.; Olckers, A.; Modiano, D.; Holmes, S.; Destro-Bisol, G. (2002). "A Back Migration from Asia to Sub-Saharan Africa is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes". The American Journal of Human Genetics. 70 (5): 1197–1214. doi:10.1086/340257. PMC 447595. PMID 11910562.
- ↑ Bosch, E.; Calafell, F.; Comas, D.; Oefner, P. J.; Underhill, P. A.; Bertranpetit, J. (2001). "High-Resolution Analysis of Human Y-Chromosome Variation Shows a Sharp Discontinuity and Limited Gene Flow between Northwestern Africa and the Iberian Peninsula". The American Journal of Human Genetics. 68 (4): 1019–29. doi:10.1086/319521. PMC 1275654. PMID 11254456.
- ↑ "Isogg 2012".
- ↑ Y Haplogroups of the World Online Edition
- ↑ Abdallah Laroui, The History of the Maghrib (Paris 1970; Princeton 1977) at 17, 60 (re S.W.Asians, referencing the earlier work of Gsell).
- ↑ Camps, Gabriel (1996), Les Berbères, Edisud, pp. 11–14, 65
- ↑ Achilli, A.; Rengo, C.; Battaglia, V.; Pala, M.; Olivieri, A.; Fornarino, S.; Magri, C.; Scozzari, R.; Babudri, N. (2005). "Saami and Berbers—An Unexpected Mitochondrial DNA Link". The American Journal of Human Genetics. 76 (5): 883–886. doi:10.1086/430073. PMC 1199377. PMID 15791543.
- ↑ J. Desanges, "The proto-Berbers" 236-245, at 237, in General History of Africa, v.II Ancient Civilizations of Africa (UNESCO 1990).
- ↑ Mário Curtis Giordani, História da África. Anterior aos descobrimentos (Petrópolis, Brasil: Editora Vozes 1985) at 42-43, 77-78. Giordani references Bousquet, Les Berbères (Paris 1961).
- 1 2 Malyarchuk, B. A.; Derenko, M.; Perkova, M.; Grzybowski, T.; Vanecek, T.; Lazur, J. (2008). "Reconstructing the phylogeny of African mitochondrial DNA lineages in Slavs". European Journal of Human Genetics. 16 (9): 1091–1096. doi:10.1038/ejhg.2008.70. PMID 18398433.
- ↑ Cherni, L.; Fernandes, V. N.; Pereira, J. B.; Costa, M. D.; Goios, A.; Frigi, S.; Yacoubi-Loueslati, B.; Amor, M. B.; Slama, A. (2009). "Post-last glacial maximum expansion from Iberia to North Africa revealed by fine characterization of mtDNA H haplogroup in Tunisia". American Journal of Physical Anthropology. 139 (2): 253–260. doi:10.1002/ajpa.20979. PMID 19090581.
- ↑ Brace, C. L. (2006). "The questionable contribution of the Neolithic and the Bronze Age to European craniofacial form". Proceedings of the National Academy of Sciences. 103: 242–247. doi:10.1073/pnas.0509801102. PMC 1325007. PMID 16371462.
- ↑ Cavalli-Sforza, L.; Menozzi, P.; Piazza, A. (1993). "Demic expansions and human evolution". Science. 259 (5095): 639–646. doi:10.1126/science.8430313. PMID 8430313.
- ↑ Bar-Yosef O (1987) Pleistocene connections between Africa and Southwest Asia: an archaeological perspective. The African Archaeological Review; Chapter 5, pg 29-38.
- ↑ Underhill, P. A.; Kivisild, T. (2007). "Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations". Annual Review of Genetics. 41: 539–564. doi:10.1146/annurev.genet.41.110306.130407. PMID 18076332.
- ↑ Kefi R, Bouzaid E, Stevanovitch A, Beraud-Colomb E. "MITOCHONDRIAL DNA AND PHYLOGENETIC ANALYSIS OF PREHISTORIC NORTH AFRICAN POPULATIONS" (PDF). ISABS. Retrieved 17 January 2016.
- ↑ Bernard Secher, Rosa Fregel, José M Larruga, Vicente M Cabrera, Phillip Endicott, José J Pestano and Ana M González. "The history of the North African mitochondrial DNA haplogroup U6 gene flow into the African, Eurasian and American continents". BMC Evolutionary Biology. Retrieved 26 March 2016.
- ↑ Fadhlaoui-Zid, K.; Plaza, S.; Calafell, F.; Ben Amor, M.; Comas, D.; Bennamar, A.; Gaaied, E. (2004). "Mitochondrial DNA Heterogeneity in Tunisian Berbers". Annals of Human Genetics. 68 (3): 222–33. doi:10.1046/j.1529-8817.2004.00096.x. PMID 15180702.
- ↑ Coudray, C.; Olivieri, A.; Achilli, A.; Pala, M.; Melhaoui, M.; Cherkaoui, M.; El-Chennawi, F.; Kossmann, M.; Torroni, A.; Dugoujon, J. M. (2009). "The Complex and Diversified Mitochondrial Gene Pool of Berber Populations". Annals of Human Genetics. 73 (2): 196–214. doi:10.1111/j.1469-1809.2008.00493.x. PMID 19053990.
- ↑ Cherni, L.; Fernandes, V. N.; Pereira, J. B.; Costa, M. D.; Goios, A.; Frigi, S.; Yacoubi-Loueslati, B.; Amor, M. B.; Slama, A. (2009). "Post-last glacial maximum expansion from Iberia to North Africa revealed by fine characterization of mtDNA H haplogroup in Tunisia". American Journal of Physical Anthropology. 139 (2): 253–260. doi:10.1002/ajpa.20979. PMID 19090581.
- ↑ Harich, N.; Costa, M. D.; Fernandes, V. N.; Kandil, M.; Pereira, J. B.; Silva, N. M.; Pereira, L. S. (2010). "The trans-Saharan slave trade - clues from interpolation analyses and high-resolution characterization of mitochondrial DNA lineages". BMC Evolutionary Biology. 10: 138. doi:10.1186/1471-2148-10-138. PMC 2875235. PMID 20459715.
- ↑ Frigi; et al. (August 2010). "Ancient Local Evolution of African mtDNA Haplogroups in Tunisian Berber Populations". Human Biology. 82 (4). doi:10.1353/hub.2010.0003.
- ↑ Esteban, E.; González-Pérez, E.; Harich, N.; López-Alomar, A.; Via, M.; Luna, F.; Moral, P. (2004). "Genetic relationships among Berbers and South Spaniards based on CD4 microsatellite/Alu haplotypes". Annals of Human Biology. 31 (2): 202–212. doi:10.1080/03014460310001652275. PMID 15204363.
- ↑ Harich et al 2010
- ↑ "Reduced Genetic Structure for Iberian Peninsula: implications for population demography. (2004)" (PDF). Retrieved 2011-08-01.
- ↑ Goncalves, R.; Freitas, A.; Branco, M.; Rosa, A.; Fernandes, A. T.; Zhivotovsky, L. A.; Underhill, P. A.; Kivisild, T.; Brehm, A. (2005). "Y-chromosome Lineages from Portugal, Madeira and Acores Record Elements of Sephardim and Berber Ancestry". Annals of Human Genetics. 69 (4): 443–54. doi:10.1111/j.1529-8817.2005.00161.x. PMID 15996172.
- ↑ Lucotte, G.; Gerard, N.; Mercier, G. (2001). "North African Genes in Iberia Studied by Y-Chromosome DNA Haplotype 5". Human Biology. 73 (5): 763–769. doi:10.1353/hub.2001.0066. PMID 11758696.
- ↑ Gérard, N.; Berriche, S.; Aouizérate, A.; Diéterlen, F.; Lucotte, G. R. (2006). "North African Berber and Arab Influences in the Western Mediterranean Revealed by Y-Chromosome DNA Haplotypes". Human Biology. 78 (3): 307–316. doi:10.1353/hub.2006.0045. PMID 17216803.
- 1 2 Cruciani, F.; La Fratta, R.; Trombetta, B.; Santolamazza, P.; Sellitto, D.; Colomb, E. B.; Dugoujon, J. -M.; Crivellaro, F.; Benincasa, T. (2007). "Tracing Past Human Male Movements in Northern/Eastern Africa and Western Eurasia: New Clues from Y-Chromosomal Haplogroups E-M78 and J-M12". Molecular Biology and Evolution. 24 (6): 1300–1311. doi:10.1093/molbev/msm049. PMID 17351267.
- ↑ Di Gaetano, C.; Cerutti, N.; Crobu, F.; Robino, C.; Inturri, S.; Gino, S.; Guarrera, S.; Underhill, P. A.; King, R. J.; et al. (2008). "Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome". European Journal of Human Genetics. 17 (1): 91–99. doi:10.1038/ejhg.2008.120. PMC 2985948. PMID 18685561. "The co-occurrence of the Berber E3b1b-M81 (2.12%) and of the Mid-Eastern J1-M267 (3.81%) Hgs together with the presence of E3b1a1-V12, E3b1a3-V22, E3b1a4-V65 (5.5%) support the hypothesis of intrusion of Moroccan genes. (...) These Hgs are common in northern Africa and are observed only in Mediterranean Europe and together the presence of the E3b1b-M81 highlights the genetic relationships between the Moroccan Almohad Empire and Sicily. (...) Hg E3b1b-M81 network cluster confirms the genetic affinity between Sicily and The Moroccan Empires."
- ↑ "The study shows that religious conversions and the subsequent marriages between people of different lineage had a relevant impact on modern populations both in Spain, especially in the Balearic Islands, and in Portugal", The religious conversions of Jews and Muslims have had a profound impact on the population of the Iberian Peninsula, Elena Bosch, 2008 Archived May 21, 2009, at the Wayback Machine.
- ↑ Capelli, C.; Onofri, V.; Brisighelli, F.; Boschi, I.; Scarnicci, F.; Masullo, M.; Ferri, G.; Tofanelli, S.; Tagliabracci, A. (2009). "Moors and Saracens in Europe: Estimating the medieval North African male legacy in southern Europe". European Journal of Human Genetics. 17 (6): 848–852. doi:10.1038/ejhg.2008.258. PMC 2947089. PMID 19156170.
- 1 2 Flores, C.; Maca-Meyer, N.; Larruga, J. M.; Cabrera, V. M.; Karadsheh, N.; Gonzalez, A. M. (2005). "Isolates in a corridor of migrations: A high-resolution analysis of Y-chromosome variation in Jordan". Journal of Human Genetics. 50 (9): 435–441. doi:10.1007/s10038-005-0274-4. PMID 16142507.
- ↑ Beleza, S.; Gusmao, L.; Lopes, A.; Alves, C.; Gomes, I.; Giouzeli, M.; Calafell, F.; Carracedo, A.; Amorim, A. (2006). "Micro-Phylogeographic and Demographic History of Portuguese Male Lineages". Annals of Human Genetics. 70 (2): 181–194. doi:10.1111/j.1529-8817.2005.00221.x. PMID 16626329.
- 1 2 Maca-Meyer, N.; Sanchez-Velasco, P.; Flores, C.; Larruga, J. -M.; Gonzalez, A. -M.; Oterino, A.; Leyva-Cobian, F. (2003). "Y Chromosome and Mitochondrial DNA Characterization of Pasiegos, a Human Isolate from Cantabria (Spain)". Annals of Human Genetics. 67 (4): 329–39. doi:10.1046/j.1469-1809.2003.00045.x. PMID 12914567.
- ↑ Fregel, R.; Gomes, V. N.; Gusmão, L.; González, A. M.; Cabrera, V. M.; Amorim, A. N.; Larruga, J. M. (2009). "Demographic history of Canary Islands male gene-pool: Replacement of native lineages by European". BMC Evolutionary Biology. 9: 181. doi:10.1186/1471-2148-9-181. PMC 2728732. PMID 19650893., see table
- ↑ See the remarks of genetic genealogist Robert Tarín for example. We can add 6.1 percent (eight out of 132) in Cuba
- ↑ Mendizabal, I.; Sandoval, K.; Berniell-Lee, G.; Calafell, F.; Salas, A.; Martinez-Fuentes, A.; Comas, D. (2008). "Genetic origin, admixture, and asymmetry in maternal and paternal human lineages in Cuba". BMC Evolutionary Biology. 8: 213. doi:10.1186/1471-2148-8-213. PMC 2492877. PMID 18644108. "The presence of chromosomes of North African origin (E3b1b-M81; Cruciani et al., 2004) can also be explained by a Portuguese-mediated influx, since this haplogroup reaches a frequency of 5.6 percent in Portugal (Beleza et al., 2006), quite similar to the frequency found in Rio de Janeiro (5.4 percent) among European contributors."
- ↑ Silva, D. A.; Carvalho, E.; Costa, G.; Tavares, L. G.; Amorim, A. N.; Gusmão, L. (2006). "Y-chromosome genetic variation in Rio De Janeiro population". American Journal of Human Biology. 18 (6): 829–837. doi:10.1002/ajhb.20567. PMID 17039481.
- ↑ Coco, C.; Magistrelli, P.; Granone, P.; Roncolini, G.; Picciocchi, A. (1992). "Conservative surgery for early cancer of the distal rectum". Diseases of the Colon & Rectum. 35 (2): 131. doi:10.1007/BF02050667.