Synaptotagmin 1
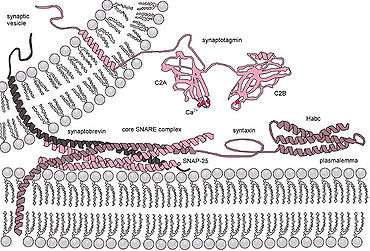
Synaptotagmin 1 (or synaptotagmin) is a Ca2+ sensor in the membrane of the pre-synaptic axon terminal, coded by gene SYT1 (previously SYT) at 12q21.2 (MIM:185605).[2]
Synaptotagmins (SYTs) constitute a family of membrane-trafficking proteins that are characterized by an N-terminal transmembrane region (TMR), a variable linker, and two C-terminal C2 domains - C2A and C2B. There are 15 members in the mammalian synaptotagmin family. There are several C2-domain containing protein families that are related to synaptotagmins, including transmembrane (Ferlins, E-Syts, and MCTPs) and soluble (RIMs, Munc13s, synaptotagmin-related proteins and B/K) proteins.
Functions
Based on their brain/endocrine distribution and biochemical properties, in particular C2 domains of certain synaptotagmins bound to calcium, synaptotagmins were proposed to function as calcium sensors in the regulation of neurotransmitter release and hormone secretion. Although synaptotagmins share a similar domain structure and a high degree of homology in the C2 domains, not all synaptotagmins bind to calcium. In fact, only eight out of the fifteen synaptotagmins are capable of calcium binding. The calcium binding synaptotagmins include synaptotagmins 1, 2, 3, 5, 6, 7, 9, and 10. The remaining seven synaptotagmins do not bind to calcium due to the lack of calcium coordinating residues or spatial orientation of the acidic residues (see the section on C2 domains for details).
Calcium-binding synaptotagmins act as Ca2+ sensors and are involved in both:
- early synaptic vesicle docking to the presynaptic membrane via interaction with β-neurexin[3] or SNAP-25[4]
- late steps of Ca2+-evoked synaptic vesicle fusion with the presynaptic membrane.[5][6][7]
It was recently shown that synaptotagmin 1 can displace complexin from the SNARE complex in the presence of calcium. This is thought to be one of the last steps in exocytosis.[8] Calcium-bound synaptotagmin binding to the SNARE complex, causes the fusion clamp effect of complexin to be released, allowing vesicle fusion to occur and exocytosis to proceed.[9]
C-terminal C2-domains
The C2 domain is a widely occurring conserved sequence motif of 130-140 amino acid residues, which was first defined as the second constant sequence in PKC isoforms. The C2 domain was first shown to bind to calcium in synaptotagmin-1. Subsequent atomic structure analysis of synaptotagmin-1 at 1.9 Å resolution indicated that its C2 domains are composed of a stable eight-stranded β-sandwich with flexible loops emerging from the top and bottom. Nuclear magnetic resonance (NMR) studies of synaptotagmin-1 revealed that calcium binds exclusively to the top loops, and the binding pockets are coordinated by five conserved aspartate residues: three calcium ions bind to C2A via D172, D178, D230, D232, S235 and D238, and two calcium ions bind to C2B via D303, D309, D363, 365 and D371. Not all synaptotagmin C2 domains bind to calcium. In fact, based on sequence similarities and subsequent confirmation by biochemical analyses, only eight synaptotagmins bind to calcium, namely, synaptotagmin-1, -2, -3, -5, -6, -7, -9 and -10. The lack of critical residues involved in calcium binding accounts for the majority of failure in calcium-binding by the other synaptotagmins. This includes both C2 domains of synaptotagmin-11, -12, -13, -14 and -15, and C2A domain of synaptotagmin-4 and -8. Synaptotagmin-4 and -11 C2B domains, which possess all five acidic residues in the top loops, however, do not bind to calcium due to spatial orientation of the calcium ligands that fail to form proper calcium binding sites. For calcium-binding synaptotagmins, although amino acid residues in the top loops other than those mentioned above are not directly involved in coordinating calcium binding, they affect calcium binding affinity, such as R233 in synaptotagmin-1. The diversity of sequences and structures flanking the calcium-coordinating amino acid residues renders the eight synaptotagmins bind to calcium at various affinities, covering the full range of calcium requirements for regulated exocytosis.
The C2A domain regulates the fusion step of synaptic vesicle exocytosis.[10][11] Consistent with this, the kinetics of Ca2+-dependent phospholipid binding activity of the C2A domain in vitro are compatible with the very fast nature of neurotransmitter release (within 200 μs).[12] The C2A domain was shown to bind negatively charged phospholipids in a Ca2+-dependent fashion. Ca2+-binding alters the protein-protein interactions of synaptotagmin such as increasing the affinity of synaptotagmin for syntaxin.
The C2B domain binds to phosphatidyl-inositol-3,4,5-triphosphate (PIP3) in the absence of calcium ions and to phosphatidylinositol bisphosphate (PIP2) in their presence, suggesting that a lipid-interaction switch occurs during depolarization. Ca2+-binding to the C2B domain confers synaptotagmin dimerization involved in the fusion step of synaptic vesicles by Ca2+-dependent self-clustering via the C2B domain. Ca2+-independent is the interaction between the C2B domain and SNAP-25, and between the C2B domain and the "synprint" (synaptic protein interaction) motif of the pore-forming subunit of voltage-gated calcium channels. The C2B domain regulates also the recycling step of synaptic vesicles by binding to the clathrin assembly protein, AP-2.
References and notes
- ↑ Georgiev, Danko D .; James F . Glazebrook (2007). "Subneuronal processing of information by solitary waves and stochastic processes". In Lyshevski, Sergey Edward. Nano and Molecular Electronics Handbook. Nano and Microengineering Series. CRC Press. pp. 17–1–17–41. ISBN 978-0-8493-8528-5.
- ↑ http://videocast.nih.gov/summary.asp?Live=11162&bhsw=1366&bhqs=1
- ↑ Fukuda M, Moreira JE, Liu V, Sugimori M, Mikoshiba K, Llinas RR (2000). "Role of the conserved WHXL motif in the C terminus of synaptotagmin in synaptic vesicle docking". Proc Natl Acad Sci USA. 97 (26): 14715–14719. doi:10.1073/pnas.260491197. PMC 18984
 . PMID 11114192.
. PMID 11114192.
- ↑ Schiavo G, Stenbeck G, Rothman JE, Söllner TH (1997). "Binding of the synaptic vesicle v-SNARE, synaptotagmin, to the plasma membrane t-SNARE, SNAP-25, can explain docked vesicles at neurotoxin-treated synapses". Proc Natl Acad Sci USA. 94 (3): 997–1001. doi:10.1073/pnas.94.3.997. PMC 19628. PMID 9023371.
- ↑ Pang ZP, Melicoff E, Padgett D, Liu Y, Teich AF, Dickey BF, et al. (2006). "Synaptotagmin-2 is essential for survival and contributes to Ca2+ triggering of neurotransmitter release in central and neuromuscular synapses". The Journal of Neuroscience. 26 (52): 13493–13504. doi:10.1523/JNEUROSCI.3519-06.2006. PMID 17192432.
- ↑ Maximov A, Südhof TC (2005). "Autonomous function of synaptotagmin 1 in triggering synchronous release independent of asynchronous release". Neuron. 48 (4): 547–554. doi:10.1016/j.neuron.2005.09.006. PMID 16301172.
- ↑ O'Connor V, Lee AG (2002). "Synaptic vesicle fusion and synaptotagmin: 2B or not 2B?". Nature Neuroscience. 5 (9): 823–824. doi:10.1038/nn0902-823. PMID 12196805.
- ↑ Tang J, Maximov A, Shin OH, Dai H, Rizo J, Südhof TC (2006). "A complexin/synaptotagmin 1 switch controls fast synaptic vesicle exocytosis". Cell. 126 (6): 1175–1187. doi:10.1016/j.cell.2006.08.030. PMID 16990140.
- ↑ Maximov, A.; Tang, J.; Yang, X.; Pang, Z. P.; Sudhof, T. C. (2009). "Complexin Controls the Force Transfer from SNARE Complexes to Membranes in Fusion". Science. 323 (5913): 516–521. doi:10.1126/science.1166505. PMC 3235366. PMID 19164751.
- ↑ Zimmerberg J, Akimov SA, Frolov V (2006). "Synaptotagmin: fusogenic role for calcium sensor?". Nature Structural & Molecular Biology. 13 (4): 301–303. doi:10.1038/nsmb0406-301. PMID 16715046.
- ↑ Fernández-Chacón R, Königstorfer A, Gerber SH, García J, Matos MF, Stevens CF, et al. (2001). "Synaptotagmin I functions as a calcium regulator of release probability". Nature. 410 (6824): 41–49. doi:10.1038/35065004. PMID 11242035.
- ↑ Chapman ER (2002). "Synaptotagmin: A Ca2+ sensor that triggers exocytosis?". Nature Reviews Molecular Cell Biology. 3 (7): 498–508. doi:10.1038/nrm855. PMID 12094216.
External links
- UMich Orientation of Proteins in Membranes families/superfamily-47 - Orientations of C2 domains in the lipid bilayer
- Synaptotagmins at the US National Library of Medicine Medical Subject Headings (MeSH)