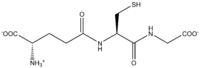
Glutathione
 | |
 | |
| Names | |
|---|---|
| IUPAC name
(2S)-2-amino-4-{[(1R)-1-[(carboxymethyl)carbamoyl]-2-sulfanylethyl]carbamoyl}butanoic acid | |
| Other names
γ-L-Glutamyl-L-cysteinylglycine (2S)-2-Amino-5-[[(2R)-1-(carboxymethylamino)-1-oxo- 3-sulfanylpropan-2-yl]amino]-5-oxopentanoic acid | |
| Identifiers | |
| 70-18-8 | |
| 3D model (Jmol) | Interactive image |
| Abbreviations | GSH |
| ChEBI | CHEBI:60836 |
| ChEMBL | ChEMBL1543 |
| ChemSpider | 111188 |
| DrugBank | DB00143 |
| ECHA InfoCard | 100.000.660 |
| 6737 | |
| KEGG | C00051 |
| MeSH | Glutathione |
| PubChem | 124886 |
| UNII | GAN16C9B8O |
| |
| |
| Properties | |
| C10H17N3O6S | |
| Molar mass | 307.32 g·mol−1 |
| Melting point | 195 °C (383 °F; 468 K) |
| Freely soluble[1] | |
| Solubility in methanol, diethyl ether | Insoluble |
| Pharmacology | |
| V03AB32 (WHO) | |
| Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
| | |
| Infobox references | |
Glutathione (GSH) is an important antioxidant in plants, animals, fungi, and some bacteria and archaea. Glutathione is capable of preventing damage to important cellular components caused by reactive oxygen species such as free radicals, peroxides, lipid peroxides, and heavy metals.[2] It is a tripeptide with a gamma peptide linkage between the carboxyl group of the glutamate side chain and the amine group of cysteine, and the carboxyl group of cysteine is attached by normal peptide linkage to a glycine.
Thiol groups are reducing agents, existing at a concentration around 5 mM in animal cells. Glutathione reduces disulfide bonds formed within cytoplasmic proteins to cysteines by serving as an electron donor. In the process, glutathione is converted to its oxidized form, glutathione disulfide (GSSG), also called L-(–)-glutathione.
Once oxidized, glutathione can be reduced back by glutathione reductase, using NADPH as an electron donor.[3] The ratio of reduced glutathione to oxidized glutathione within cells is often used as a measure of cellular oxidative stress.[4][5]
Biosynthesis
The biosynthesis pathway for glutathione is found in some bacteria, such as cyanobacteria and proteobacteria, but is missing in many other bacteria. Most eukaryotes, including humans, synthesize glutathione, but some do not, such as Leguminosae, Entamoeba, and Giardia. The only archaea that make glutathione are halobacteria.[6][7]
Glutathione is not an essential nutrient for humans, since it can be synthesized in the body from the amino acids L-cysteine, L-glutamic acid, and glycine; it does not have to be present as a supplement in the diet. The sulfhydryl group (SH) of cysteine serves as a proton donor and is responsible for its biological activity. Cysteine is the rate-limiting factor in cellular glutathione biosynthesis, since this amino acid is relatively rare in foods.
Cells make glutathione in two adenosine triphosphate-dependent steps:
- First, gamma-glutamylcysteine is synthesized from L-glutamate and cysteine via the enzyme gamma-glutamylcysteine synthetase (glutamate cysteine ligase, GCL). This reaction is the rate-limiting step in glutathione synthesis.[8]
- Second, glycine is added to the C-terminal of gamma-glutamylcysteine via the enzyme glutathione synthetase.
Animal glutamate cysteine ligase (GCL) is a heterodimeric enzyme composed of a catalytic and a modulatory subunit. The catalytic subunit is necessary and sufficient for all GCL enzymatic activity, whereas the modulatory subunit increases the catalytic efficiency of the enzyme. Mice lacking the catalytic subunit (i.e., lacking all de novo GSH synthesis) die before birth.[9] Mice lacking the modulatory subunit demonstrate no obvious phenotype, but exhibit marked decrease in GSH and increased sensitivity to toxic insults.[10][11][12]
While all animal cells are capable of synthesizing glutathione, glutathione synthesis in the liver has been shown to be essential. Mice with genetically induced loss of GCLC (i.e., GSH synthesis) only in the liver die within a month of birth.[13][14] Major transport into the blood stream is driven by an electrochemical gradient, specifically through the transport proteins RcGshT and RsGshT.[15] Similarly, bile is a medium in which GSH and GSSG is exported to.[14][16]
The plant glutamate cysteine ligase (GCL) is a redox-sensitive homodimeric enzyme, conserved in the plant kingdom.[17] In an oxidizing environment, intermolecular disulfide bridges are formed and the enzyme switches to the dimeric active state. The midpoint potential of the critical cysteine pair is -318 mV. In addition to the redox-dependent control, the plant GCL enzyme is feedback inhibited by glutathione.[18] GCL is exclusively located in plastids, and glutathione synthetasei(GS)s dual-targeted to plastids and cytosol, thus GSH and gamma-glutamylcysteine are exported from the plastids.[19] Both glutathione biosynthesis enzymes are essential in plants; knock-outs of GCL and GS are lethal to embryo and seedling.[20]
Function
Glutathione exists in both reduced (GSH) and oxidized (GSSG) states. In the reduced state, the thiol group of cysteine is able to donate a reducing equivalent (H++ e−) to other molecules, such as reactive oxygen species to neutralize them, or to protein cysteines to maintain their reduced forms. With donating an electron, glutathione itself becomes reactive and readily reacts with another reactive glutathione to form glutathione disulfide (GSSG). Such a reaction is probable due to the relatively high concentration of glutathione in cells (up to 7 mM in the liver).[21]
Generally, interactions between GSH and other molecules with higher relative electrophilicity deplete GSH levels within the cell. An exception to this case involves the sensitivity of GSH to the electrophilic compound's relative concentration. In high concentrations, the organic molecule Diethyl maleate fully depleted GSH levels in cells. However, in low concentrations, a minor decrease in cellular GSH levels was followed by a two-fold increase.[22][23]
GSH can be regenerated from GSSG by the enzyme glutathione reductase (GSR):[3] NADPH reduces FAD present in GSR to produce a transient FADH-anion. This anion then quickly breaks a disulfide bond (Cys58 - Cys63) and leads to Cys63's nucleophilically attacking the nearest sulfide unit in the GSSG molecule (promoted by His467), which creates a mixed disulfide bond (GS-Cys58) and a GS-anion. His467 of GSR then protonates the GS-anion to form the first GSH. Next, Cys63 nucleophilically attacks the sulfide of Cys58, releasing a GS-anion, which, in turn, picks up a solvent proton and is released from the enzyme, thereby creating the second GSH. So, for every GSSG and NADPH, two reduced GSH molecules are gained, which can again act as antioxidants scavenging reactive oxygen species in the cell.
In healthy cells and tissue, more than 90% of the total glutathione pool is in the reduced form (GSH) and less than 10% exists in the disulfide form (GSSG). An increased GSSG-to-GSH ratio is considered indicative of oxidative stress.[24]
Glutathione has multiple functions:
- It maintains levels of reduced glutaredoxin and glutathione peroxidase[25]
- It is one of the major endogenous antioxidant produced by the cells, participating directly in the neutralization of free radicals and reactive oxygen compounds, as well as maintaining exogenous antioxidants such as vitamins C and E in their reduced (active) forms.[26][27][28]
- Regulation of the nitric oxide cycle is critical for life, but can be problematic if unregulated.[29]
- It is used in metabolic and biochemical reactions such as DNA synthesis and repair, protein synthesis, prostaglandin synthesis, amino acid transport, and enzyme activation. Thus, every system in the body can be affected by the state of the glutathione system, especially the immune system, the nervous system, the gastrointestinal system, and the lungs.
- It has a vital function in iron metabolism. Yeast cells depleted of GSH or containing toxic levels of GSH show an intense iron starvation-like response and impairment of the activity of extramitochondrial ISC enzymes thus inhibiting oxidative endoplasmic reticulum folding, followed by death.[30]
- It has roles in progression of the cell cycle, including cell death.[5] GSH levels regulate redox changes to nuclear proteins necessary for the initiation of cell differentiation. Differences in GSH levels also determine the expressed mode of cell death, being either apoptosis or cell necrosis. Manageably low levels result in the systematic breakage of the cell whereas excessively low levels result in rapid cell death.[31]
Function in animals
GSH is known as a substrate in conjugation reactions, which is catalyzed by glutathione S-transferase enzymes in cytosol, microsomes, and mitochondria. However, GSH is also capable of participating in nonenzymatic conjugation with some chemicals.
In the case of N-acetyl-p-benzoquinone imine (NAPQI), the reactive cytochrome P450-reactive metabolite formed by paracetamol (acetaminophen), which becomes toxic when GSH is depleted by an overdose of acetaminophen, glutathione is an essential antidote to overdose. Glutathione conjugates to NAPQI and helps to detoxify it. In this capacity, it protects cellular protein thiol groups, which would otherwise become covalently modified; when all GSH has been spent, NAPQI begins to react with the cellular proteins, killing the cells in the process. The preferred treatment for an overdose of this painkiller is the administration (usually in atomized form) of N-acetyl-L-cysteine (often as a preparation called Mucomyst[32]), which is processed by cells to L-cysteine and used in the de novo synthesis of GSH.
Glutathione (GSH) participates in leukotriene synthesis and is a cofactor for the enzyme glutathione peroxidase. It is also important as a hydrophilic molecule that is added to lipophilic toxins and waste in the liver during biotransformation before they can become part of the bile. Glutathione is also needed for the detoxification of methylglyoxal, a toxin produced as a byproduct of metabolism.
This detoxification reaction is carried out by the glyoxalase system. Glyoxalase I (EC 4.4.1.5) catalyzes the conversion of methylglyoxal and reduced glutathione to S-D-lactoyl-glutathione. Glyoxalase II (EC 3.1.2.6) catalyzes the hydrolysis of S-D-lactoyl-glutathione to glutathione and D-lactic acid.
Glutathione, along with oxidized glutathione (GSSG) and S-nitrosoglutathione (GSNO), have been found to bind to the glutamate recognition site of the NMDA and AMPA receptors (via their γ-glutamyl moieties), and may be endogenous neuromodulators.[33][34][35] At millimolar concentrations, they may also modulate the redox state of the NMDA receptor complex.[34] In addition, glutathione has been found to bind to and activate ionotropic receptors that are different from any other excitatory amino acid receptor, and which may constitute glutathione receptors, potentially making it a neurotransmitter.[36]
Function in plants
In plants, glutathione is crucial for biotic and abiotic stress management. It is a pivotal component of the glutathione-ascorbate cycle, a system that reduces poisonous hydrogen peroxide.[37] It is the precursor of phytochelatins, glutathione oligomers that chelate heavy metals such as cadmium.[38] Glutathione is required for efficient defence against plant pathogens such as Pseudomonas syringae and Phytophthora brassicae.[39] Adenylyl-sulfate reductase, an enzyme of the sulfur assimilation pathway, uses glutathione as an electron donor. Other enzymes using glutathione as a substrate are glutaredoxins. These small oxidoreductases are involved in flower development, salicylic acid, and plant defence signalling.[40]
Supplementation
Calcitriol (1,25-dihydroxyvitamin D3), the active metabolite of vitamin D3, after being synthesized from calcifediol in the kidney, increases glutathione levels in the brain and appears to be a catalyst for glutathione production.[41] It takes about ten days for the body to process vitamin D3 into calcitriol.[42]
S-adenosylmethionine (SAMe), a cosubstrate involved in methyl group transfer, has also been shown to increase cellular glutathione content in persons suffering from a disease-related glutathione deficiency.[43][44][45]
Low glutathione is commonly observed in wasting and negative nitrogen balance, as seen in cancer, HIV/AIDS, sepsis, trauma, burns, and athletic overtraining. Low levels are also observed in periods of starvation. These effects are hypothesized to be influenced by the higher glycolytic activity associated with cachexia, which result from reduced levels of oxidative phosphorylation.[46][47]
Bioavailability
Glutathione is only to a small extent bioavailable to humans; the human body is capable of maintaining a consistent level of GSH. Oral introduction of GSH into the body is, in fact, scarcely effective to increase its plasma and/or intracellular concentration. At the base of its poor bioavailability is the nature of glutathione which, being a tripeptide, is the substrate of proteases (peptidases) of the alimentary canal, and the absence of a specific carrier of glutathione at the level of cell membrane.[48][49]
Methods to determine glutathione
Small molecule based glutathione probes
Ellman's reagent and Monobromobimane
Reduced glutathione may be visualized using Ellman's reagent or bimane derivatives such as monobromobimane. The monobromobimane method is more sensitive. In this procedure, cells are lysed and thiols extracted using a HCl buffer. The thiols are then reduced with dithiothreitol and labelled by monobromobimane. Monobromobimane becomes fluorescent after binding to GSH. The thiols are then separated by HPLC and the fluorescence quantified with a fluorescence detector.
Monochlorobimane
Monochlorobimane can be used to quantify glutathione in vivo. The quantification is done by confocal laser scanning microscopy after application of the dye to living cells.[50] This quantification process relies on measuring the rates of fluorescence changes and is limited to plant cells.
5-Chloromethylfluorescein diacetate (CMFDA)
CMFDA was initially used as a cell tracker. Unfortunately, it has also been mistakenly used as a glutathione probe. Unlike monochlorobimane, whose fluorescence increases upon reacting with glutathione, the fluorescence increase of CMFDA is due to the hydrolysis of the acetate groups inside cells. Although CMFDA may react with glutathione in cells, the fluorescence increase does not reflect the reaction. Therefore, studies using CMFDA as a glutathione probe should be revisited and re-interpreted.[51][52]
ThiolQuant Green
The major limitation of these bimane based probes and many other reported probes is that these probes are based on irreversible chemical reactions with glutathione, which renders these probes incapable of monitoring the real-time glutathione dynamics. Recently, the first reversible reaction based fluorescent probe-ThiolQuant Green (TQG)-for glutathione was reported.[53] ThiolQuant Green can not only perform high resolution measurements of glutathione levels in single cells using a confocal microscope, but also be applied in flow cytometry to perform bulk measurements.
Protein based glutathione probes
Another approach, which allows to measure the glutathione redox potential at a high spatial and temporal resolution in living cells is based on redox imaging using the redox-sensitive green fluorescent protein (roGFP)[54] or redox sensitive yellow fluorescent protein (rxYFP)[55] GSSG because its very low physiological concentration is difficult to measure accurately unless the procedure is carefully executed and monitored and the occurrence of interfering compounds is properly addressed. GSSG concentration ranges from 10 to 50 μM in all solid tissues, and from 2 to 5 μM in blood (13–33 nmol per gram Hb). GSH-to-GSSG ratio ranges from 100 to 700.[56]
Other biological implications
Cancer
Once a tumor has been established, elevated levels of glutathione may act to protect cancerous cells by conferring resistance to chemotherapeutic drugs.[57] The antineoplastic mustard drug canfosfamide was modelled on the structure of glutathione.
Cystic fibrosis
Several studies have been completed on the effectiveness of introducing inhaled glutathione to people with cystic fibrosis with mixed results.[58][59]
Alzheimer's disease
Whilst extracellular Aβ plaques, NFT, inflammation in the form or reactive astrocytes and microglia, and neuronal loss are all consistent pathological features of AD, a mechanistic link between these factors is yet to be clarified. Although the majority of past research has focused on fibrillar Aβ, soluble oligomeric Aβ species are now considered to be of major pathological importance in AD. Up-regulation of GSH may be protective against the oxidative and neurotoxic effects of oligomeric Aβ.
Uses
Winemaking
The content of glutathione in must, the first raw form of wine, determines the browning, or caramelizing effect, during the production of white wine by trapping the caffeoyltartaric acid quinones generated by enzymic oxidation as grape reaction product.[60] Its concentration in wine can be determined by mass spectrometry.[61]
Cosmetics
Glutathione plays an important role in preventing oxidative damage to the skin.[62] In addition to its many recognized biological functions, glutathione has also been associated with skin lightening ability.[63] The role of glutathione as a skin whitener was discovered as a side effect of large doses of glutathione.[64] Glutathione utilizes different mechanisms to exert its action as a skin whitening agent at various levels of melanogenesis. It inhibits melanin synthesis by means of stopping the neurotransmitter precursor L-DOPA’s ability to interact with tyrosinase in the process of melanin production.[65] Glutathione inhibits the actual production as well as agglutination of melanin by interrupting the function of L-DOPA. Another study found that glutathione inhibits melanin formation by direct inactivation of the enzyme tyrosinase by binding and chelating copper within the enzyme’s active site.[66] Glutathione’s antioxidant property allows it to inhibit melanin synthesis by quenching of free radicals and peroxides that contribute to tyrosinase activation and melanin formation.[67] Its antioxidant property also protects the skin from UV radiation and other environmental as well as internal stressors that generate free radicals that cause skin damage and hyperpigmentation.[68] In most mammals, melanin formation consists of eumelanin (brown-black pigment) and pheomelanin ( yellow-red pigment) as either mixtures or co-polymers.[69] Increase in glutathione level may induce the pigment cell to produce pheomelanin instead of eumelanin pigments.[70] A research by Te-Sheng Chang found lowest levels of reduced glutathione to be associated with eumelanin type pigmentation, whereas the highest ones were associated with the pheomelanin.[63] As a result, it is reasonable to assume that depletion of glutathione would result in eumelanin formation. Prota [71] observed that decreased glutathione concentration led to the conversion of L-Dopaquinone to Dopachrome, increasing the formation of brown-black pigment (eumelanin).
Importance of gamma-glutamylcysteine as a precursor for glutathione synthesis
Gamma-glutamylcysteine (GGC) is the immediate precursor to GSH. GGC supplementation would circumvent feedback inhibitory control of GCL by the end product GSH. Accordingly, a method of elevating GSH levels with the notable advantage of bypassing negative feedback inhibition has been described. Because of this, GGC has been the focus of therapeutic efforts since Puri and Meister 1983. The first documented use of GGC in brains appears to be Pileblad and Magnusson, 1992. Astroglia cells are capable of utilising GGC.[72] Direct delivery of the GSH precursor GCC to brain has been reported to effectively replenish levels of GSH in the brain.[73]
Most of the work done on GGC has been preclinical, based on in vivo animal models, or in vitro brain cultures. In order for the therapeutic value of GGC elevation against AD to be vindicated, three empirical hurdles have to be cleared. The first is to demonstrate that delivery of GCC into the brain can indeed increase GSH.[73] The second is to demonstrate that the increase in GGC can indeed reduce oxidative stress in the brain,[74] a condition frequently linked with cognitive decline.
See also
- Glutathione synthetase deficiency
- Ophthalmic acid
- roGFP, a tool to measure the cellular glutathione redox potential
- Glutathione-ascorbate cycle
- Bacterial glutathione transferase
- Thioredoxin, a cysteine-containing small proteins with very similar functions as reducing agents
- Glutaredoxin, an antioxidant protein that uses reduced glutathione as a cofactor and is reduced nonenzymatically by it
- Bacillithiol
- Mycothiol
- gamma-L-Glutamyl-L-cysteine
References
- 1 2 Merck Index, 11th Edition, 4369
- ↑ Pompella, A; Visvikis, A; Paolicchi, A; Tata, V; Casini, AF (October 2003). "The changing faces of glutathione, a cellular protagonist". Biochemical Pharmacology. 66 (8): 1499–503. doi:10.1016/S0006-2952(03)00504-5. PMID 14555227.
- 1 2 Couto, Narciso; Malys, Naglis; Gaskell, Simon; Barber, Jill (30 April 2013). "Partition and Turnover of Glutathione Reductase from Saccharomyces cerevisiae: a Proteomic Approach". Journal of Proteome Research. 12 (6): 2885–94. doi:10.1021/pr4001948. PMID 23631642.
- ↑ Pastore, Anna; Piemonte, Fiorella; Locatelli, Mattia; Lo Russo, Anna Lo; Gaeta, Laura Maria; Tozzi, Giulia; Federici, Giorgio (August 2001). "Determination of blood total, reduced, and oxidized glutathione in pediatric subjects". Clinical Chemistry. 47 (8): 1467–9. PMID 11468240.
- 1 2 Lu, Shelly C. (1 May 2013). "Glutathione synthesis". Biochimica et Biophysica Acta. 1830 (5): 3143–53. doi:10.1016/j.bbagen.2012.09.008. ISSN 0006-3002. PMC 3549305
 . PMID 22995213. (subscription required (help)).
. PMID 22995213. (subscription required (help)). - ↑ Copley, Shelley D; Dhillon, Jasvinder K (29 April 2002). "Lateral gene transfer and parallel evolution in the history of glutathione biosynthesis genes". Genome Biology. 3 (5): research0025.1. doi:10.1186/gb-2002-3-5-research0025. PMC 115227. PMID 12049666.
- ↑ Wonisch, Willibald; Schaur, Rudolf J. (2001). "Chapter 2: Chemistry of Glutathione". In Grill, D.; Tausz, T.; De Kok, L.J. Significance of glutathione in plant adaptation to the environment. Springer. ISBN 1-4020-0178-9 – via Google Books.
- ↑ White, C. C.; Viernes, H.; Krejsa, C. M.; Botta, D.; Kavanagh, T. J. (15 July 2003). "Fluorescence-based microtiter plate assay for glutamate–cysteine ligase activity". Analytical Biochemistry. 318 (2): 175–80. doi:10.1016/S0003-2697(03)00143-X. PMID 12814619. (subscription required (help)).
- ↑ Dalton, T; Dieter, MZ; Yang, Y; Shertzer, HG; Nebert, DW (20 December 2000). "Knockout of the Mouse Glutamate Cysteine Ligase Catalytic Subunit (Gclc) Gene: Embryonic Lethal When Homozygous, and Proposed Model for Moderate Glutathione Deficiency When Heterozygous". Biochemical and Biophysical Research Communications. 279 (2): 324–9. doi:10.1006/bbrc.2000.3930. PMID 11118286. (subscription required (help)).
- ↑ Yang, Y.; Dieter, MZ; Chen, Y; Shertzer, HG; Nebert, DW; Dalton, TP (2002). "Initial characterization of the glutamate-cysteine ligase modifier subunit Gclm(-/-) knockout mouse. Novel model system for a severely compromised oxidative stress response". Journal of Biological Chemistry. 277 (51): 49446–52. doi:10.1074/jbc.M209372200. PMID 12384496.
- ↑ Giordano, G; Afsharinejad, Z; Guizzetti, M; Vitalone, A; Kavanagh, T; Costa, L (2007). "Organophosphorus insecticides chlorpyrifos and diazinon and oxidative stress in neuronal cells in a genetic model of glutathione deficiency". Toxicology and Applied Pharmacology. 219 (2–3): 181–9. doi:10.1016/j.taap.2006.09.016. PMID 17084875.
- ↑ McConnachie, L. A.; Mohar, I.; Hudson, F. N.; Ware, C. B.; Ladiges, W. C.; Fernandez, C.; Chatterton-Kirchmeier, S.; White, C. C.; Pierce, R. H.; Kavanagh, T. J. (2007). "Glutamate Cysteine Ligase Modifier Subunit Deficiency and Gender as Determinants of Acetaminophen-Induced Hepatotoxicity in Mice". Toxicological Sciences. 99 (2): 628–36. doi:10.1093/toxsci/kfm165. PMID 17584759.
- ↑ Chen, Ying; Yang, Yi; Miller, Marian L.; Shen, Dongxiao; Shertzer, Howard G.; Stringer, Keith F.; Wang, Bin; Schneider, Scott N.; Nebert, Daniel W.; Dalton, Timothy P. (2007). "Hepatocyte-specificGclcdeletion leads to rapid onset of steatosis with mitochondrial injury and liver failure". Hepatology. 45 (5): 1118–28. doi:10.1002/hep.21635. PMID 17464988.
- 1 2 Sies, Helmut (1999-11-01). "Glutathione and its role in cellular functions". Free Radical Biology and Medicine. 27 (9–10): 916–921. doi:10.1016/S0891-5849(99)00177-X.
- ↑ Li, L.; Lee, T. K.; Ballatori, N. (1997-08-01). "Functional re-evaluation of the putative glutathione transporters, RcGshT and RsGshT". The Yale Journal of Biology and Medicine. 70 (4): 301–310. ISSN 0044-0086. PMC 2589333. PMID 9626750.
- ↑ Lee, T. K.; Li, L.; Ballatori, N. (1997-08-01). "Hepatic glutathione and glutathione S-conjugate transport mechanisms". The Yale Journal of Biology and Medicine. 70 (4): 287–300. ISSN 0044-0086. PMC 2589341. PMID 9626749.
- ↑ Hothorn, M.; Wachter, A; Gromes, R; Stuwe, T; Rausch, T; Scheffzek, K (2006). "Structural Basis for the Redox Control of Plant Glutamate Cysteine Ligase". Journal of Biological Chemistry. 281 (37): 27557–65. doi:10.1074/jbc.M602770200. PMID 16766527.
- ↑ Hicks, L. M.; Cahoon, R. E.; Bonner, E. R.; Rivard, R. S.; Sheffield, J.; Jez, J. M. (2007). "Thiol-Based Regulation of Redox-Active Glutamate-Cysteine Ligase from Arabidopsis thaliana". The Plant Cell Online. 19 (8): 2653–61. doi:10.1105/tpc.107.052597. PMC 2002632. PMID 17766407.
- ↑ Wachter, Andreas; Wolf, Sebastian; Steininger, Heike; Bogs, Jochen; Rausch, Thomas (2004). "Differential targeting of GSH1 and GSH2 is achieved by multiple transcription initiation: implications for the compartmentation of glutathione biosynthesis in the Brassicaceae". The Plant Journal. 41 (1): 15–30. doi:10.1111/j.1365-313X.2004.02269.x. PMID 15610346.
- ↑ Pasternak, Maciej; Lim, Benson; Wirtz, Markus; Hell, RüDiger; Cobbett, Christopher S.; Meyer, Andreas J. (2007). "Restricting glutathione biosynthesis to the cytosol is sufficient for normal plant development". The Plant Journal. 53 (6): 999–1012. doi:10.1111/j.1365-313X.2007.03389.x. PMID 18088327.
- ↑ Kaplowitz, N. (1981-01-01). "The importance and regulation of hepatic glutathione.". The Yale Journal of Biology and Medicine. 54 (6): 497–502. ISSN 0044-0086. PMC 2596047. PMID 7342494.
- ↑ Bannai, Shiro; Tateishi, Noriko (1986-02-01). "Role of membrane transport in metabolism and function of glutathione in mammals". The Journal of Membrane Biology. 89 (1): 1–8. doi:10.1007/BF01870891. ISSN 0022-2631.
- ↑ Bannai, S. (1984-02-25). "Induction of cystine and glutamate transport activity in human fibroblasts by diethyl maleate and other electrophilic agents". The Journal of Biological Chemistry. 259 (4): 2435–2440. ISSN 0021-9258. PMID 6142042.
- ↑ Halprin, Kenneth (1967). "The Measurement of Glutathione in Human Epidermis using Glutathione Reductase". Journal of Investigative Dermatology. 48 (2): 149. doi:10.1038/jid.1967.24.
- ↑ Grant, Chris M. (2001-02-01). "Role of the glutathione/glutaredoxin and thioredoxin systems in yeast growth and response to stress conditions". Molecular Microbiology. 39 (3): 533–541. doi:10.1046/j.1365-2958.2001.02283.x. ISSN 1365-2958.
- ↑ Dringen, R. (2000-12-01). "Metabolism and functions of glutathione in brain". Progress in Neurobiology. 62 (6): 649–671. doi:10.1016/s0301-0082(99)00060-x. ISSN 0301-0082. PMID 10880854.
- ↑ Scholz, RW. Graham KS. Gumpricht E. Reddy CC. (1989). "Mechanism of interaction of vitamin E and glutathione in the protection against membrane lipid peroxidation". Ann NY Acad Sci. 570: 514–7. doi:10.1111/j.1749-6632.1989.tb14973.x.
- ↑ Hughes, RE (1964). "Reduction of dehydroascorbic acid by animal tissues". Nature. 203 (4949): 1068–9. doi:10.1038/2031068a0.
- ↑ Clementi, Emilio; Smith, Guy Charles; Howden, Martin; Dietrich, Salvador; Bugg, S; O'Connell, MJ; Goldsbrough, PB; Cobbett, CS (1999). "Phytochelatin synthase genes from Arabidopsis and the yeast Schizosaccharomyces pombe". The Plant cell. 11 (6): 1153–64. doi:10.1105/tpc.11.6.1153. JSTOR 3870806. PMC 144235. PMID 10368185.
- ↑ Kumar, Chitranshu; et al. (2011). "Glutathione revisited: a vital function in iron metabolism and ancillary role in thiol-redox control". The EMBO Journal. 30: 2044–2056. doi:10.1038/emboj.2011.105.
- ↑ Hall, A. G. (1999-03-01). "Review: The role of glutathione in the regulation of apoptosis". European Journal of Clinical Investigation. 29 (3): 238–245. doi:10.1046/j.1365-2362.1999.00447.x. ISSN 0014-2972. PMID 10202381.
- ↑ "Pharmaceutical Information - MUCOMYST". RxMed. Retrieved 2014-02-13.
- ↑ Steullet, P.; Neijt, H.C.; Cuénod, M.; Do, K.Q. (2006). "Synaptic plasticity impairment and hypofunction of NMDA receptors induced by glutathione deficit: Relevance to schizophrenia". Neuroscience. 137 (3): 807–819. doi:10.1016/j.neuroscience.2005.10.014. ISSN 0306-4522.
- 1 2 Varga, V.; Jenei, Zs.; Janáky, R.; Saransaari, P.; Oja, S. S. (1997). "Glutathione Is an Endogenous Ligand of Rat Brain N-Methyl-D-Aspartate (NMDA) and 2-Amino-3-Hydroxy-5-Methyl-4-Isoxazolepropionate (AMPA) Receptors". Neurochemical Research. 22 (9): 1165–1171. doi:10.1023/A:1027377605054. ISSN 0364-3190.
- ↑ Janáky, R.; Ogita, K.; Pasqualotto, B. A.; Bains, J. S.; Oja, S. S.; Yoneda, Y.; Shaw, C. A. (1999-09-01). "Glutathione and signal transduction in the mammalian CNS". Journal of Neurochemistry. 73 (3): 889–902. ISSN 0022-3042. PMID 10461878.
- ↑ Oja, S (2000). "Modulation of glutamate receptor functions by glutathione". Neurochemistry International. 37 (2-3): 299–306. doi:10.1016/S0197-0186(00)00031-0. ISSN 0197-0186.
- ↑ Noctor, Graham; Foyer, Christine H. (1998). "ASCORBATE AND GLUTATHIONE: Keeping Active Oxygen Under Control". Annual Review of Plant Physiology and Plant Molecular Biology. 49 (1): 249–279. doi:10.1146/annurev.arplant.49.1.249. PMID 15012235.
- ↑ Ha, S.-B.; Smith, AP; Howden, R; Dietrich, WM; Bugg, S; O'Connell, MJ; Goldsbrough, PB; Cobbett, CS (1999). "Phytochelatin Synthase Genes from Arabidopsis and the Yeast Schizosaccharomyces pombe". The Plant Cell Online. 11 (6): 1153–64. doi:10.1105/tpc.11.6.1153. PMC 144235. PMID 10368185.
- ↑ Parisy, Vincent; Poinssot, Benoit; Owsianowski, Lucas; Buchala, Antony; Glazebrook, Jane; Mauch, Felix (2006). "Identification of PAD2 as a γ-glutamylcysteine synthetase highlights the importance of glutathione in disease resistance of Arabidopsis". The Plant Journal. 49 (1): 159–72. doi:10.1111/j.1365-313X.2006.02938.x. PMID 17144898.
- ↑ Rouhier, Nicolas; Lemaire, StéPhane D.; Jacquot, Jean-Pierre (2008). "The Role of Glutathione in Photosynthetic Organisms: Emerging Functions for Glutaredoxins and Glutathionylation". Annual Review of Plant Biology. 59 (1): 143–66. doi:10.1146/annurev.arplant.59.032607.092811. PMID 18444899.
- ↑ Garcion, E; Wion-Barbot, N; Montero-Menei, C; Berger, F; Wion, D (2002). "New clues about vitamin D functions in the nervous system". Trends in Endocrinology and Metabolism. 13 (3): 100–5. doi:10.1016/S1043-2760(01)00547-1. PMID 11893522.
- ↑ Van Groningen, L.; Opdenoordt, S.; Van Sorge, A.; Telting, D.; Giesen, A.; De Boer, H. (2010). "Cholecalciferol loading dose guideline for vitamin D-deficient adults". European Journal of Endocrinology. 162 (4): 805–811. doi:10.1530/EJE-09-0932. PMID 20139241.
- ↑ Lieber, Charles S. (2002). "S-adenosyl-L-methionine: its role in the treatment of liver disorders". The American Journal of Clinical Nutrition. 76 (5): 1183S–7S. PMID 12418503.
- ↑ Vendemiale, G.; Altomare, E.; Trizio, T.; Le Grazie, C.; Di Padova, C.; Salerno, M. T.; Carrieri, V.; Albano, O. (1989). "Effects of Oral S-Adenosyl-l-Methionine on Hepatic Glutathione in Patients with Liver Disease". Scandinavian Journal of Gastroenterology. 24 (4): 407–15. doi:10.3109/00365528909093067. PMID 2781235.
- ↑ Loguercio, C; Nardi, G; Argenzio, F; Aurilio, C; Petrone, E; Grella, A; Del Vecchio Blanco, C; Coltorti, M (1994). "Effect of S-adenosyl-L-methionine administration on red blood cell cysteine and glutathione levels in alcoholic patients with and without liver disease". Alcohol and alcoholism (Oxford, Oxfordshire). 29 (5): 597–604. PMID 7811344.
- ↑ Dröge, Wulf; Holm, Eggert (1997). "Role of cysteine and glutathione in HIV infection and other diseases associated with muscle wasting and immunological dysfunction". The FASEB Journal. 11 (13): 1077–89. PMID 9367343.
- ↑ Tateishi, Noriko; Higashi, Taneaki; Shinya, Shintaro; Naruse, Akiko; Sakamoto, Yukiya (1974-01-01). "Studies on the Regulation of Glutathione Level in Rat Liver". Journal of Biochemistry. 75 (1): 93–103. ISSN 0021-924X. PMID 4151174.
- ↑ Allen, J; Bradley, RD (Sep 2011). "Effects of oral glutathione supplementation on systemic oxidative stress biomarkers in human volunteers". J Altern Complement Med. 17 (9): 827–33. doi:10.1089/acm.2010.0716.
- ↑ Witschi, A; Reddy, S; Stofer, B; Lauterburg, BH (1992). "The systemic availability of oral glutathione". Eur J Clin Pharmacol. 43 (6): 667–9. doi:10.1007/bf02284971.
- ↑ Meyer, Andreas J.; May, Mike J.; Fricker, Mark (2001). "Quantitative in vivo measurement of glutathione in Arabidopsis cells". The Plant Journal. 27 (1): 67–78. doi:10.1046/j.1365-313x.2001.01071.x. PMID 11489184.
- ↑ Sebastià, Jordi; Cristòfol, Rosa; Martín, Manuela; Rodríguez-Farré, Eduard; Sanfeliu, Coral (2003-01-01). "Evaluation of fluorescent dyes for measuring intracellular glutathione content in primary cultures of human neurons and neuroblastoma SH-SY5Y". Cytometry Part A. 51 (1): 16–25. doi:10.1002/cyto.a.10003. ISSN 1552-4922. PMID 12500301.
- ↑ Lantz, R. Clark; Lemus, Ranulfo; Lange, Robert W.; Karol, Meryl H. (2001-04-01). "Rapid Reduction of Intracellular Glutathione in Human Bronchial Epithelial Cells Exposed to Occupational Levels of Toluene Diisocyanate". Toxicological Sciences. 60 (2): 348–355. doi:10.1093/toxsci/60.2.348. ISSN 1096-6080. PMID 11248147.
- ↑ Jiang, Xiqian; Yu, Yong; Chen, Jianwei; Zhao, Mingkun; Chen, Hui; Song, Xianzhou; Matzuk, Alexander J.; Carroll, Shaina L.; Tan, Xiao; Sizovs, Antons; Cheng, Ninghui; Wang, Meng C.; Wang, Jin (2015). "Quantitative Imaging of Glutathione in Live Cells Using a Reversible Reaction-Based Ratiometric Fluorescent Probe". ACS Chemical Biology. 10: 864–874. doi:10.1021/cb500986w.
- ↑ Meyer, Andreas J.; Brach, Thorsten; Marty, Laurent; Kreye, Susanne; Rouhier, Nicolas; Jacquot, Jean-Pierre; Hell, RüDiger (2007). "Redox-sensitive GFP inArabidopsis thalianais a quantitative biosensor for the redox potential of the cellular glutathione redox buffer". The Plant Journal. 52 (5): 973–86. doi:10.1111/j.1365-313X.2007.03280.x. PMID 17892447.
- ↑ Maulucci, Giuseppe; Labate, Valentina; Mele, Marina; Panieri, Emiliano; Arcovito, Giuseppe; Galeotti, Tommaso; Østergaard, H; Winther, JR; De Spirito, Marco; Pani, G. (2008). "High-resolution imaging of redox signaling in live cells through an oxidation-sensitive yellow fluorescent protein". Science Signaling. 1 (43): pl3. doi:10.1126/scisignal.143pl3. PMID 18957692.
- ↑ Giustarini, D; Dalle-Donne, I; Milzani, A; Fanti, P; Rossi, R (Sep 2013). "Analysis of GSH and GSSG after derivatization with N-ethylmaleimide". Nat Protoc. 8 (9): 1660–9. doi:10.1038/nprot.2013.095. PMID 23928499.
- ↑ Balendiran, Ganesaratnam K.; Dabur, Rajesh; Fraser, Deborah (2004). "The role of glutathione in cancer". Cell Biochemistry and Function. 22 (6): 343–52. doi:10.1002/cbf.1149. PMID 15386533.
- ↑ Visca A, Bishop CT, Hilton SC, Hudson VM. "Improvement in clinical markers in CF patients using a reduced glutathione regimen: an uncontrolled, observational study. J Cyst Fibros 2008
- ↑ Bishop, C; Hudson, VM; Hilton, SC; Wilde, C (2005). "A pilot study of the effect of inhaled buffered reduced glutathione on the clinical status of patients with cystic fibrosis". Chest. 127: 308–317. doi:10.1378/chest.127.1.308.
- ↑ Rigaud, Jacques; Cheynier, Véronique; Souquet, Jean-Marc; Moutounet, Michel (1991). "Influence of must composition on phenolic oxidation kinetics". Journal of the Science of Food and Agriculture. 57 (1): 55–63. doi:10.1002/jsfa.2740570107.
- ↑ Vallverdú-Queralt, Anna; Verbaere, Arnaud; Meudec, Emmanuelle; Cheynier, Veronique; Sommerer, Nicolas (2015). "Straightforward Method To Quantify GSH, GSSG, GRP, and Hydroxycinnamic Acids in Wines by UPLC-MRM-MS.". J. Agric. Food Chem. 63: 142–149. doi:10.1021/jf504383g.
- ↑ Jansen, A. H.; Russell, B. J.; Chernick, V (1975). "Respiratory effects of H+ and dinitrophenol injections into the brain stem subarachnoid space of fetal lambs". Canadian Journal of Physiology and Pharmacology. 53 (5): 726–33. doi:10.1139/y75-101. PMID 134.
- 1 2 Libíková, H; Pogády, J; Wiedermann, V; Breier, S (1975). "Search for herpetic antibodies in the cerebrospinal fluid in senile dementia and mental retardation". Acta virologica. 19 (6): 493–5. PMC 2443094. PMID 1996.
- ↑ Prasad, Sahdeo; Srivastava, Smita; Singh, Madhulika; Shukla, Yogeshwer (2009). "Clastogenic Effects of Glyphosate in Bone Marrow Cells of Swiss Albino Mice". Journal of Toxicology. 2009: 1–6. doi:10.1155/2009/308985.
- ↑ Matsuki, M; Watanabe, T; Ogasawara, A; Mikami, T; Matsumoto, T (2008). "Inhibitory mechanism of melanin synthesis by glutathione". Yakugaku Zasshi. 128 (8): 1203–7. doi:10.1248/yakushi.128.1203. PMID 18670186.
- ↑ Scott, D. M.; Mazurkiewicz, M; Leeman, P (1976). "The long-term monitoring of ventilation rhythms of the polychaetous annelid Nereis virens sars". Comparative Biochemistry and Physiology A. 53 (1): 65–8. doi:10.1016/s0300-9629(76)80012-6. PMID 187.
- ↑ Karg, E; Odh, G; Wittbjer, A; Rosengren, E; Rorsman, H (1993). "Hydrogen peroxide as an inducer of elevated tyrosinase level in melanoma cells". The Journal of Investigative Dermatology. 100 (2 Suppl): 209S–213S. doi:10.1111/1523-1747.ep12465218. PMID 8433009.
- ↑ Shindo, Y; Hashimoto, T (1995). "Antioxidant defence mechanism of the skin against UV irradiation: Study of the role of catalase using acatalasaemia fibroblasts". Archives of dermatological research. 287 (8): 747–53. doi:10.1007/bf01105800. PMID 8554387.
- ↑ Ito, S (1993). "High-performance liquid chromatography (HPLC) analysis of eu- and pheomelanin in melanogenesis control". The Journal of Investigative Dermatology. 100 (2 Suppl): 166S–171S. doi:10.1038/jid.1993.71. PMID 8433004.
- ↑ Jara, J. R.; Aroca, P; Solano, F; Martinez, J. H.; Lozano, J. A. (1988). "The role of sulfhydryl compounds in mammalian melanogenesis: The effect of cysteine and glutathione upon tyrosinase and the intermediates of the pathway". Biochimica et Biophysica Acta. 967 (2): 296–303. doi:10.1016/0304-4165(88)90023-2. PMID 2903772.
- ↑ Tan, A. W.; Nuttall, F. Q. (1975). "Characteristics of the dephosphorylated form of phosphorylase purified from rat liver and measurement of its activity in crude liver preparations". Biochimica et Biophysica Acta. 410 (1): 45–60. doi:10.1016/0005-2744(75)90206-5. PMID 75.
- ↑ Dringen, R (1997). "Use of Dipeptides for the Synthesis of Glutathione by Astroglia-Rich Primary Cultures". Journal of Neurochemistry. 69 (2): 868–74. PMID 9231749.
- 1 2 Pileblad, E (1992). "Increase in rat brain glutathione following intracerebroventricular administration of gamma-glutamylcysteine". Biochemical Pharmacology. 44 (5): 895–903. doi:10.1016/0006-2952(92)90121-x. PMID 1530658.
- ↑ Le, TM (2011). "Glutamylcysteine ameliorates oxidative injury in neurons and astrocytes in vitro and increases brain glutathione in vivo". Neurotoxicology. 32 (5): 518–525. doi:10.1016/j.neuro.2010.11.008. PMC 3079792. PMID 21159318.
Related research
- Bilinsky, LM; Reed, MC; Nijhout, HF (2015). "The role of skeletal muscle in liver glutathione metabolism during acetaminophen overdose". Journal of Theoretical Bology. 376: 118–33. doi:10.1016/j.jtbi.2015.04.006. PMID 25890031. Lay summary – ALN Magazine (24 June 2015).

- Drevet, JR (2006). "The antioxidant glutathione peroxidase family and spermatozoa: A complex story". Molecular and Cellular Endocrinology. 250 (1–2): 70–9. doi:10.1016/j.mce.2005.12.027. PMID 16427183.

- Wu, Guoyao; Fang, Yun-Zhong; Yang, Sheng; Lupton, Joanne R.; Turner, Nancy D. (2004). "Glutathione metabolism and its implications for health". The Journal of Nutrition. 134 (3): 489–92. PMID 14988435.
External links
- Glutathione bound to proteins in the PDB