Mongoloid








Mongoloid /ˈmɒŋ.ɡə.lɔɪd/[1][2] is a term for peoples indigenous to East Asia, Central Asia, Southeast Asia, Siberia or North Asia, Northern Europe, the Arctic, the Americas,parts of the Pacific Islands, parts of Africa (Madagascar), and South Asia. Individuals within these populations often share certain associated phenotypic traits, such as epicanthic folds (epicanthus), sinodonty, and neoteny.
Epicanthic folds and oblique palpebral fissures are common among most Mongoloid individuals. Most exhibit Mongolian spot from birth to about age four.[3][4] The vast majority have straight, black hair and dark brown almond-shaped eyes, and most have broad, relatively flat faces as well.[5]
Traits more controversially ascribed to Mongoloids include high intelligence[6][7][8] and a stoic, taciturn, unaggressive demeanor. The extent to which individual psychological makeup is a social construct, rather than a matter of biology, remains a matter of debate. (See also the articles Race and intelligence and Model minority.)
The term "mongoloid" was introduced by 18th century ethnologists to describe Central Asian and East Asian populations, as part of a tripartite typological model of race: Mongoloid, Caucasoid, and Negroid. Although some forensic anthropologists and other scientists continue to use such terms in some contexts (such as criminal justice), their usage is now discouraged by most anthropologists due to the questionable nature of such models. In addition, "Mongoloid" has had a second usage, now generally avoided as highly offensive: until the late 20th century, people with Down syndrome[9][10][11][12] were often referred to as "Mongoloids" or in terms of "Mongolian idiocy" or "Mongolian imbecility".
Geographic range and populations included



Mongoloid or Asiatic peoples are the most spread out among all human populations since they have stretched almost completely around the earth's surface. They can be found as far "east" as Greenland, and yet also as far to the "west" as Kalmykia, the Crimea, and even Scandinavia,[17] giving Asiatic peoples or their descendants a historical presence across the vast expanse of four continents. According to the Meyers Konversations-Lexikon (1885–90), peoples included in the Mongoloid race are North Mongol, Chinese & Indochinese, Japanese & Korean, Tibetan, Malay, Polynesian, Maori, Micronesian, Eskimo, and Native American.[18]
In 1856, the "Mongolian" race, using a narrow definition which did not include either the "Malay" or the "American" races, was the second most populous race in the world behind the Caucasian race.[19] In 1881, the Mongoloid race, using a broad definition which included both Malays and indigenous Americans, was the most populous race on Earth,[20] and it was still the most populous race on Earth in the year 1892, using a narrow definition which did not include either the "Malayan" or the "American" races.[21] In 1994, the Mongoloid race, using a broad definition which included indigenous Americans, comprised 34% of the Earth's human population which made it the second most populous race behind Caucasoids who comprised 56% of the Earth's human population at that time.[22]
In addition, "Mongoloid" has had a second usage, now generally avoided as highly offensive: until the late 20th century, people with Down syndrome[9][10][11][12] were often referred to as "Mongoloids" or in terms of "Mongolian idiocy" or "Mongolian imbecility".
The first use of the term Mongolian race was by Christoph Meiners in 1785, who divided humanity into two races he labeled "Tartar-Caucasians" and "Mongolians".[23]
Johann Friedrich Blumenbach said that he borrowed the term Mongolian from Christoph Meiners to describe the race he designated "second, [which] includes that part of Asia beyond the Ganges and below the river Amoor, which looks toward the south, together with the islands and the greater part of these countries which is now called Australian".[24]
In 1861, Isidore Geoffroy Saint-Hilaire added the Australian as a secondary race (subrace) of the principal race of Mongolian.[25] In the nineteenth century Georges Cuvier used the term Mongolian again as a racial classification, but additionally included American Indians under the term.[26] Arthur de Gobineau defined the extent of the Mongolian race, "by the yellow the Altaic, Mongol, Finnish and Tartar branches".[27][28] Later, Thomas Huxley used the term Mongoloid and included American Indians as well as Arctic Native Americans.[29] Other terms were proposed, such as Mesochroi (middle color),[30] but Mongoloid was widely adopted.
In 1940, anthropologist Franz Boas included the American race as part of the Mongoloid race of which he mentioned the Aztecs of Mexico and the Maya of Yucatan.[31] Boas also said that, out of the races of the Old World, the American native had features most similar to the east Asiatic.[31]
In 1981, Elizabeth Smithgall Watts who taught anthropology at Tulane University[32] said that the question of American Indians being a separate race from "Asiatic Mongoloids" is a question of how much genetic difference a population needs from another population to be considered a "major race". She said that even the people who consider American Indians to be a separate race acknowledge that they are genetically closest to "Asians".[33]
In 1983, Douglas J. Futuyma, professor of evolutionary processes at the University of Michigan, said that the inclusion of Native Americans and Pacific Islanders under the Mongoloid race was not recognized by many anthropologists who consider them distinct races.[34]
In 1984, Roger J. Lederer, Professor of Biological Sciences at California State University at Chico,[35] separately listed the Mongoloid race from Pacific islanders and American Indians when he enumerated the "geographical variants of the same species known as races...we recognize several races, Inuit, American Indians, Mongoloid... Polynesian".[36]
In 1995, Dr. Marta Mirazon Lahr of the Department of Biological Anthropology at Cambridge University used the term Mongoloid to refer to Asian populations, Indigenous Australians, Pacific Islanders, Negritos, and Amerindians, classifying Northeast Asians as typical Mongoloids and all other Mongoloid groups as atypical Mongoloids.[37]
Finns were previously considered by some scholars to be partly Mongoloid, dating to claims by Johann Friedrich Blumenbach. Finns (and other Finno-Ugrians in Europe) are now considered typically European.[38] 10% of Finnish genes are shared with Siberian populations. Nevertheless more than 80% of Finnish genes are from a single ancient Northeastern European population.[39]
The Sami people of the Finno-Ugric brance are also included as Mongoloid as of their genes, origin and physical appearance,[17] although in recent times many of the indigenous people has been inflicted with the Nordic genes through mixing of the local population between the Scandinavian people and the aboriginals in the Sapmi region.123
Subraces
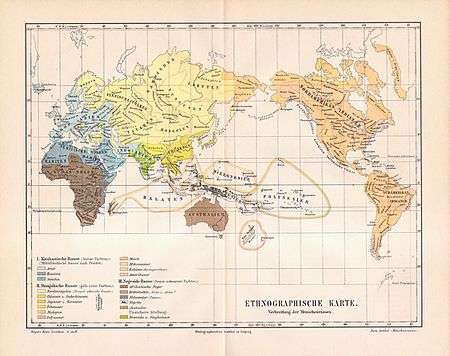
| Caucasoid race: Negroid race: Uncertain: | Mongoloid race: |
In 1900, Joseph Deniker said the "Mongol race admits two varieties or subraces: Tunguse or Northern Mongolian... and Southern Mongolian".[25]
Archaeologist Peter Bellwood claims that the vast majority of people in Southeast Asia, the region he calls the "clinal Mongoloid-Australoid zone", are Southern Mongoloids but have a high degree of Australoid admixture.[40]
Professor of anthropology, Akazawa Takeru (赤沢威) at the International Research Center for Japanese Studies, Kyoto, said that there are Neo-Mongoloids and Paleo-Mongoloids. Akazawa said Neo-Mongoloids have "extreme Mongoloid, cold-adapted features" and they include the Chinese, Buryats, Eskimo and Chukchi. In contrast, Akazawa said Paleo-Mongoloids are less cold-adapted. He said Burmese, Filipinos, Polynesians, Jōmon and the indigenous peoples of the Americas were Paleo-Mongoloid.[41]
Human skeletal remains in Southeast Asia show the gradual replacement of the indigenous Australo-Melanesians by Southern Mongoloids from Southern China. No skeletal remains in Southeast Asia dated to the Pleistocene epoch have been unearthed that would classified as being indisputably Mongoloid, although skeletal remains dated to this epoch have been found with Mongoloid traits. Skeletal remains in Southeast Asia dated to the Pleistocene epoch with Mongoloid traits indicate that Mongoloid admixture from areas north of Southeast Asia was already taking place at this time. This trend toward an increasingly Mongoloid skeletal character in Southeast Asia continued during the later Holocene epoch as an increasing number of the skeletal remains dated to the last 7,000 years are classified as having "Southern Mongoloid skeletal material" relative to the earlier epochs. The dental evidence that pre-historic Southeast Asian skeletal remains are of the sundadont dental type, and the dental evidence that Southeast Asians, including Negritos, are of the sundadont dental type supports the idea that it was sundadont Southern Mongoloids from Southern China whose gene flow was making Southeast Asia more Mongoloid instead of the sinodont Northeast Asian Mongoloids from farther north. Most of the Southern Mongoloids' gradual replacement of the indigenous Australo-Melanesians in Southeast Asia, a process done by "replacing Australo-Melanesian hunter-gatherers or assimilating populations of 'Proto-Malays'", was done "within the historical period". After the "gradual and complex replacement" of the indigenous Australo-Melanesians by Southern Mongoloids in Southeast Asia, the only remaining indigenous Australo-Melanesian population in Southeast Asia at the present time are the Negritos of Thailand, Malaysia, the Philippines and the Andaman Islands. The concept which is "[t]he important concept" here is that the gradual replacement of Australo-Melanesians by Southern Mongoloids in Southeast Asia was a gradual change in the cline between these two populations.[42]
History of the concept
The earliest systematic use of the term was by Blumenbach in De generis humani varietate nativa (On the Natural Variety of Mankind, University of Göttingen, first published in 1775, re-issued with alteration of the title-page in 1776). Blumenbach included East and South East Asians, but not Native Americans or Malays, who were each assigned separate categories.
In 1865, biologist Thomas Huxley presented the views of polygenesists (Huxley was not one of them) as "some imagine their assumed species of mankind were created where we find them... the Mongolians from the Orangs".[43]
In 1964, archaeologist Kwang-chih Chang said that it seemed like the Mongoloid race originated in South China, and he said that it seemed like the Mongoloid race was differentiating itself from other races in the Late Pleistocene. Chang based these thoughts on a skull found in Sichuan and a skull found in Guangxi.[44]
In 1972, physical anthropologist Carleton Coon said, "From a hyborean [sic] group there evolved, in northern Asia, the ancestral strain of the entire specialized Mongoloid family".[45] In 1962, Coon believed that the Mongoloid "subspecies" existed "during most of the Pleistocene, from 500,000 to 10,000 years ago".[46] According to Coon, the Mongoloid race had not completed its "invasions and expansions" into Southeast Asia, the Americas, and the Pacific Islands until "[t]oward the end of the Pleistocene".[46] By this time Coon hypothesis that the Mongoloid race had become "sapien".[46]
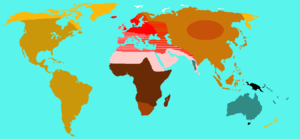
Mahinder Kumar Bhasin (Hindi: महेंद्र कुमार भसीन) of the Department of Anthropology at the University of Delhi suggested in a review of an article referencing Mourant 1983 that "The Caucasoids and the Mongoloid almost certainly became differentiated from one another somewhere in Asia" and that "Another differentiation, which probably took place in Asia, is that of the Australoids, perhaps from a common type before the separation of the Mongoloids".[48]
Paleo-anthropologist Milford Wolpoff and Rachel Caspari characterize "his [Carleton Coon's] contention [as being] that the Mongoloid race crossed the 'sapiens threshold' first and thereby evolved the furthest".[49]
Douglas J. Futuyma, professor of evolutionary processes at the University of Michigan, said the Mongoloid race "diverged 41,000 years ago" from a Mongoloid and Caucasoid group which diverged from Negroids "110,000 years ago".[34]
In 1996, professor of anthropology, Akazawa Takeru of the International Research Center for Japanese Studies, Kyoto, said Mongoloids originated in Xinjiang during the "Ice Age".[41]

In 1999, Peter Brown of the Department of Anthropology and Paleoanthropology at the University of New England evaluated three sites with early East Asian modern human skeletal remains (Liujiang, Liuzhou, Guangxi, China; Shandingdong Man of (but not Peking Man) Zhoukoudian's Upper Cave; and Minatogawa in Okinawa) dated to between 10,175 to 33,200 years ago, and finds lack of support for the conventional designation of skeletons from this period as "Proto-Mongoloid". He stated that "The colonisation of the Americas by 11 kyr indicates an earlier date for the appearance of distinctively East Asian features, however, the earliest unequivocal evidence for anatomically East Asian people on the Asian mainland remains at 7000 years BP." He saw this as "possibility that migration across the Bering Strait went in two directions and the first morphological Mongoloids evolved in the Americas." [50]
In 2006, Yali Xue (Chinese: 薛雅丽) et al. of the genome research Sanger Institute conducted a study of linkage disequilibrium that said that northern populations in East Asia started to expand in number between 34 and 22 thousand years ago, before the last glacial maximum at 21–18 KYA, while southern populations started to expand between 18 and 12 KYA, but then grew faster, and suggests that the northern populations expanded earlier because they could exploit the abundant megafauna of the "Mammoth Steppe", while the southern populations could increase in number only when a warmer and more stable climate led to more plentiful plant resources such as tubers.[51]
Features
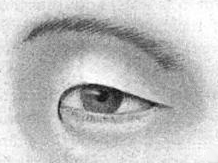
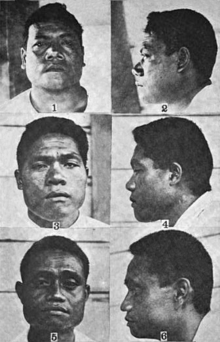

There are "many Mongoloid groups" that characteristically have "enormously broad ascending ramus".[53]
Palatine tori and mandibular tori are more commonly present in Mongoloids than in Caucasians and Negroids.[54]
A "mandibular torus" is a trait that commonly occurs in "Mongoloid populations".[55]
Australoids have teeth that are "Megadont", both Negroids and Mongoloids have teeth that are "Mesodont", and Caucasians have teeth that are "Microdont".[56]
In "Whites" and in "Mongoloid populations", the shafts of the femurs curve toward the front of the person relative to how the femurs are in "Blacks".[57]
"Mongoloids" have femurs with more curvature and more twisting at the neck than the femurs of both "whites" and "blacks". Whites have femurs that are "intermediate in both curvature and twisting" between Mongoloids and blacks. Blacks have femurs with less curvature and less twisting at the head or neck than the femurs of both whites and Mongoloids.[58]
In 1962, Carleton S. Coon said that one of the reasons that Mongoloids have flatter faces than Caucasoids is due to the masseter and temporalis jaw muscles in the faces of Mongoloids being positioned more toward the front of the faces of Mongoloids relative to where these jaw muscles are positioned in the faces of Caucasoids.[59]
A 1992 study compared the features of North African skull samples dated to the Late Pleistocene against purported "mongoloid" and "australoid" features. The study found that the skull samples had at "moderate to high frequencies" the "Chinese features" of shovel-shaped incisors and a horizontally flat face, and the study found that the skull samples had at "moderate to high frequencies" the "southeast Asian traits" of a high degree of prognathism, strong brow ridges, projecting cheekbones and "malar tuberosities".[60]
According to George W. Gill and other modern forensic anthropologists, physical traits of Mongoloid crania are generally distinct from those of the Caucasoid and Negroid races. They assert that they can identify a Mongoloid skull with an accuracy of up to 95%.[61] However, Alan H. Goodman cautions that this precision estimate is often based on methodologies using subsets of samples. He also argues that scientists have a professional and ethical duty to avoid such biological analyses since they could potentially have sociopolitical effects.[62]
Variation in craniofacial form between humans has been found to be largely due to differing patterns of biological inheritance. Modern cross-analysis of osteological variables and genome-wide SNPs has identified specific genes, which control this craniofacial development. Of these genes, DCHS2, RUNX2, GLI3, PAX1 and PAX3 were found to determine nasal morphology, whereas EDAR impacts chin protrusion.[63]
Mongoloid teeth are larger than Caucasoid and Negroid teeth. Mongoloids have mandibles that are "robust", and Mongoloids have mandibles that are "similar" to the mandibles of Negroids in respect to the chins of Mongoloids and Negroids not being as prominent as the chins of Caucasoids and in respect to the chins of Mongoloids and Negroids being "median" while the Caucasoid chin is "bilateral".[64]
Mongoloids generally have big incisors, big canines, big molars and small premolars.[65]
The East Polynesian, the Paleoindian/North American Archaic, and the Mongoloid/Late Amerindian are characterized by a "Square, heavy jaw". The East Polynesian and the Mongoloid/Late Amerindian are characterized by a "Median chin". The European is characterized by a "sharp, thin jaw" that has a "strong, prominent chin". Mongoloid peoples, meaning modern East Asians and Amerindians of the later time periods, are characterized by "robust" cheekbones that project forward and to either side of the face.[66]
The nasal sill bones of American Indians are of medium development and "sometimes even sharp", and, in this respect, they are like the nasal sill bones of "Whites" whose nasal sill bones are almost without exception sharp. The nasal bones of East Asians are "small" and "often flat". American Indians and East Asians almost never have a nasion depression which is the depression between the brow ridge and the bridge of the nose. The nasal sill bones of East Polynesians are "rounded", smooth and "dull" and, in this respect, they are like the nasal sill bones of sub-Saharan Africans and Australians/Melanesians. The nasal bones of East Polynesians are "large and prominent" and there is often a nasion depression in East Polynesians which is a trait that is also present in "Whites". East Polynesians have a lower nasal root than "Europeans". The nasal bridge of East Polynesians is not as straight in profile as the "European" nasal bridge, and the nasal bridge of East Polynesians does not have the "steeple shape" of the "Caucasoid" nasal bridge.[66]
Mongoloids have the smallest eyeballs, Negroids have eyeballs of medium size, and Caucasoids have the largest eyeballs. This eyeball size pattern is different for the size of the eye sockets in which the eyeballs reside, however, since Negroids have the smallest eye sockets, Caucasoids have eye sockets of medium size, and Mongoloids have the largest eye sockets.[67]
The Mongoloid skull shows a round head shape with a medium-width nasal aperture, rounded orbital margins, massive cheekbones, weak or absent canine fossae, moderate prognathism, absent brow ridges, simple cranial sutures, prominent zygomatic bones, broad, flat, tented nasal root, short nasal spine, shovel-shaped upper incisor teeth (scooped out behind), straight nasal profile, moderately wide palate shape, arched sagittal contour, wide facial breadth and a flatter face.— Caroline Wilkinson, Forensic Facial Reconstruction. (2004). page 86.[67]
Mongoloid subjects had about "20% higher bone density at the angle of the mandible" when compared to Caucasoid subjects.[68]
Samoans are of the Mongoloid race but their features represent a "slightly different evolution since the time of their separation and isolation from their parental stock" or a retention of features that have been lost in other Mongoloid types. The "straight" or "low waves" hair of the Samoan is one such retention compared to the stiff, coarse hair that typifies the Mongoloid. Most of the characteristics of the Samoan have Mongoloid affinities such as: skin color, hair color, eye color, conjuctiva, amount of beard, hair on chest, nasal bridge, nostrils, lips, face width, biogonial width, cephalo-facial index, nasal height, ear height and chin. Polynesians lack characteristic Mongoloid shovel-shaped incisors, because this characteristic Mongoloid trait disappeared in the Polynesian population as the teeth of Polynesians reduced in size over the course of their evolutionary history.[69]
Mongoloid features are a mesocranic skull, fairly large and protruding cheekbones, nasal bones that are flat and broad, a nasal bridge that is slightly concave without depression in the nasion, "the lower borders of the piriform aperture are not sharp but guttered", shallow prenasal fossae, small anterior nasal spine, trace amounts of canine fossae and moderate alveolar prognathism.[70]
The Paleoindian has proto-Mongoloid morphology such as pronounced development of supraorbital ridges low frontals, marked post-orbital constriction, prominent and protruding occipitals, small mastoids, long crania and a relatively narrow bizygomatic breadth.[37]
The Mongoloid eyelid is characterized by puffiness of the upper eyelid, "superficial expansion of the levator aponeurosis" that are "turned up around this transverse ligament to become the orbital septum", "low position of the preaponeurotic fat" and narrowness of the palpebral fissure.[71]
The Mongoloid racial type is distinguished by forward-projecting malar (cheek) bones, comparatively flat faces, large circular orbits, "moderate nasal aperture with a slightly pointed lower margin", larger, more gracile braincase, broader skull, broader face and flatter roof of the nose.[72]
Mongoloid skin has thick skin cuticle and an abundance of carotene (yellow pigment).[41] Mongoloid males have "little or no facial or body hair".[73] Mongoloid hair is coarse, straight, blue-black and weighs the most out of the races.[74] The size of the average Mongoloid hair is 0.0051 square millimetres (7.9×10−6 sq in) based on samples from Chinese, North and South American Indians, Eskimos and Thais.[75] Mongoloid hair whether it be Sioux, Ifugao or Japanese has the thickest diameter out of all human hair.[76] Mongoloids evolved hairlessness to keep clean while wearing heavy garments for months without bathing during the Ice Age.[41]
In 1996, Rebecca Haydenblit of the Hominid Evolutionary Biology Research Group at Cambridge University did a study on the dentition of four pre-Columbian Mesoamerican populations and compared their data to other Mongoloid populations.[77] She said that Tlatilco, Cuicuilco, Monte Albán and Cholula populations followed an overall Sundadont dental pattern characteristic of Southeast Asia rather than a Sinodont dental pattern characteristic of Northeast Asia.[77]
The traits of the Mongoloid skull are: long and broad skulls of intermediate height, arched sagittal contour, very wide facial contour, high face height, rounded orbital opening, narrow nasal opening, wide, flat nasal bones, sharp lower nasal margin, straight facial profile, moderate and white palate shape, 90%+ shovel-shaped incisors and large, smooth general form.[78]
Miquel Hernández of the Department of Animal Biology at the University of Barcelona said East Asians (Kyushu, Atayal, Philippines, Chinese, Hokkaido and Anyang) and Amerinds (Yaujos, Santa Cruz and Arikara) have the typical Mongoloid cranial pattern, but other Mongoloids such as Pacific groups (Easter Island, Mokapu, Guam and Moriori people), arctic groups (Eskimos and Buriats), Fuegians (Selk’nam, Ya´mana, Kawe´skar) and the Ainu differ from this by having "larger cranial dimensions over many variables".[79]
Commenting on the lack of body hair (glabrousness) of Negroids and Mongoloids, Carleton S. Coon wrote that "[b]oth negroid and mongoloid skin conditions are inimical to excessive hair development except upon the scalp."[80]
The theoretical index of hair bending stiffness is calculated using the thickest and thinnest axial diameters of human hair, and this index differs by race. The hair stiffness indexes of Mongoloids, Africans and Europeans are: 4.23, 2.75 and 1.59, respectively. This means that Mongoloids with the highest hair stiffness index value of 4.23 have the most rigid hair and Europeans with the lowest hair stiffness index value of 1.59 have the least rigid hair. The eccentricity of hair cross-sectional shape index is also calculated using the thickest and thinnest axial diameters of human hair, and this index also differs by race. The hair eccentricity indexes of Africans, Europeans and Mongoloids are: 1.74, 1.49 and 1.30, respectively. This means that Africans with the highest hair eccentricity index value of 1.74 have the curliest hair and Mongoloids with the lowest hair eccentricity index value of 1.30 have the least curly hair.[81]
| Mongoloid | Caucasoid | Negroid | |
|---|---|---|---|
| Cranial form | broad | medium | long |
| Sagittal outline | high, globular | high, rounded | highly variable, post-bregmatic depression |
| Nose form | medium | narrow | broad |
| Nasal bone size | small | large | medium/small |
| Nasal profile | concave | straight | straight/concave |
| Nasal spine | medium | prominent, straight | reduced |
| Nasal sill | medium | sharp | dull/absent |
| Incisor form | shoveled | blade | blade |
| Mongoloid | Caucasoid | Negroid | |
| Facial prognathism | moderate | reduced | prominent |
| Alveolar prognathism | moderate | reduced | prominent |
| Malar form | projecting | reduced | reduced |
| Palatal form | parabolic/elliptic | parabolic | hyperbolic |
| Orbital form | round | rhomboid | round |
| Mandible | robust | medium | gracile, oblique gonial angle |
| Chin projection | moderate | prominent | reduced |
| Chin form | median | bilateral | median |
Proto-Mongoloids

Mark J. Hudson, Professor of Anthropology at Nishikyushu University, Kanzaki, Saga, Japan, said Japan was settled by a Proto-Mongoloid population in the Pleistocene who became the Jōmon and their features can be seen in the Ainu and Okinawan people. Hudson said that, later, during the Yayoi period, the Neo-Mongoloid type entered Japan. Hudson said that genetically Japanese people are primarily Neo-Mongoloid with Proto-Mongoloid admixture.[83]

Theodore G. Schurr of the Department of Anthropology at University of Pennsylvania said that Mongoloid traits emerged from Transbaikalia, central and eastern regions of Mongolia, and several regions of Northern China. Schurr said that studies of cranio-facial variation in Mongolia suggest that the region of modern-day Mongolians is the origin of the Mongoloid racial type".[72]
Dr. Rukang Wu (Chinese: 吴汝康) of the Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica, China, said that the remains of Liukiang human fossils were an early type of evolving Mongoloid that indicated South China was the birthplace where the Mongoloid race originated.[70]
Dr. Marta Mirazon Lahr of the Department of Biological Anthropology at Cambridge University said there are two hypotheses on the origin of Mongoloids. Lahr said that one hypothesis is that Mongoloids originated in north Asia due to the regional continuity in this region and this population conforming best to the standard Mongoloid features. Lahr said that the other hypothesis is that Mongoloids originate from Southeast Asian populations that expanded from Africa to Southeast Asia during the first half of the Upper Pleistocene and then traveled to Australia-Melanesia and East Asia. Lahr said that the morphology of the Paleoindian is consistent with the proto-Mongoloid definition.[37]
Neoteny

Ashley Montagu claimed "the skeleton of the classic Mongoloid type is very delicately made, even down to the character of the sutures of the skull which, like those of the infant skull, are relatively smooth and untortuous. In fact the Mongoloid presents so many physical traits which are associated with the late fetus or young infant that he has been called a fetalized, infantilized or pedomorphic type. Those who have carefully observed young babies may recall that the root of the nose is frequently flat or low as in Mongoloids, and that an internal epicanthic fold in such instances is usually present. The smaller number of individual head hairs and the marked hairlessness of the remainder of the body are infantile traits, as are likewise the small mastoid processes, the shallow fossa into which the jawbone fits (the mandibular fossa), the rather stocky build, the large brain-pan and brain, lack of brow ridges, and quite a number of other characters."[5]
Stephen Oppenheimer of the Institute of Cognitive & Evolutionary Anthropology at Oxford University said, "An interesting hypothesis put forward by paleontologist Stephen Jay Gould many years ago was that the package of the Mongoloid anatomical changes could be explained by the phenomenon of neoteny, whereby an infantile or childlike body form is preserved in adult life. Neoteny in hominids is still one of the simplest explanations of how we developed a disproportionately large brain so rapidly over the past few million years. The relatively large brain and the forward rotation of the skull on the spinal column, and body hair loss, both characteristic of humans, are found in foetal chimps. Gould suggested a mild intensification of neoteny in Mongoloids, in whom it has been given the name pedomorphy. Such a mechanism is likely to involve only a few controller genes and could therefore happen over a relatively short evolutionary period. It would also explain how the counterintuitive retrousse [turned up at the end] nose and relative loss of facial hair got into the package". "[D]ecrease unnecessary muscle bulk, less tooth mass, thinner bones and smaller physical size; ...this follows the selective adaptive model of Mongoloid evolution".[85]

Paul Storm of the Naturalis Biodiversity Center, Netherlands, said that in Australasia there are two types of cranial morphologies—the "Sunda" (Mongoloid) and "Sahul" (Australoid) types. Storm said that the "Sunda" (Mongoloid) type includes Chinese and Javanese people, and he said that the "Sahul" (Australoid) type includes Papuans and Australian aborigines. Storm said that the "Sunda" (Mongoloid) type has a flat face with high cheek bones, and Storm said that this "flat face" of the Chinese and Javanese is known as the "mongoloid face". Storm further said that the "Sunda" (Mongoloid) type has a more rounded skull, "feminine (juvenile) characters", a "retention of juvenile characters" and a limited outgrowth of superstructures such as the supraorbital region. Storm said that "Sunda" (Mongoloid) skulls resemble female skulls more than "Sahul" (Australoid) skulls resemble female skulls. Storm said that the skulls of "Asian" males ("Chinese and Javanese") have "more feminine characteristics", and he said that they have "many feminine characters in contrast with Australians".[86]

Paul Storm said that Asia contained humans with "generalized" cranial morphology, but between 20,000 BP and 12,000 BP this generalized type disappeared as a new type emerged. This new type had a flatter face with more pronounced cheekbones, a more rounded head, reduced sexual dimorphism (male skulls started to resemble female skulls), a reduction of superstructures such as the supraorbital region and an increased "retention of juvenile characters". Storm said that this new type of skull that emerged is called the "Proto-Sunda" (Proto-Mongoloid) type, and it is distinguished from the "Sunda" (Mongoloid) type by being more "robust". Storm said that the "Mongoloid" or "Asian" type of skull developed relatively fast during a population bottleneck in Asia that happened during the Late Pleistocene or Early Holocene through a microevolutionary trend that involved a "continuation of neoteny and gracilisation trends". Due to different courses of evolution, Storm said that these two types of skulls, the "Sunda" (Mongoloid) type and the "Sahul" (Australoid) type, are now clearly recognizable at the present time.[86]
Andrew Arthur Abbie who was an anatomist and anthropologist at the University of Adelaide[87] talked about leg-to-torso length being related to neoteny. Abbie said that women normally have shorter legs than men, and he said that shorter legs are the normal condition in some ethnic groups such as Mongoloids. Abbie said that Mongoloids of whom he listed the people of "China, Japan and the Americas" have proportionately larger heads and shorter legs than Europeans, and he said that this is a case of "paedomorphism". Abbie said that aboriginal Australians and some African ethnic groups such as the "Negro", the "Hottentot" and the "Nubian" peoples have proportionately longer legs than Europeans, and he said that this is a case of "gerontomorphism". Abbie said that ethnic groups with proportionately shorter legs than Europeans are relatively "paedomorphic" in terms of leg-to-torso ratios when compared to Europeans, and he said that ethnic groups with proportionately longer legs than Europeans are relatively "gerontomorphic" in terms of leg-to-torso ratios when compared to Europeans.[88]
Cold adaptation


Professor of anthropology, Akazawa Takeru of the International Research Center for Japanese Studies in Kyoto Japan, said that Mongoloid features are an adaptation to the cold of the Mammoth steppe.[41] He mentions the Lewis waves of warm blood cyclical vasodilation and vasoconstriction of the peripheral capillaries in Mongoloids as an adaption to the cold.[41] He lists the short limbs, short noses, flat faces, epicanthic fold and lower surface to mass ratio as further Mongoloid adaptations to cold.[41]
Nicholas Wade said that biologists have speculated that the Mongoloid skull type was the result of natural selection in response to a cold climate, and Wade said that the Mongoloid skull type first started to indisputably appear in the archaeological record 10,000 years ago. Wade said that biologists have speculated that the fat in the eyelids of Mongoloids and the stocky builds of Mongoloids were selected for as adaptations to the cold.[89]
Takasaki Yuji (高崎裕治) of Akita University, Japan,[90] in the Journal of Physiological Anthropology and Applied Human Science said that, "Mongoloid ancestors had evolved over time in cold environments" and the short limbs of the Mongoloid was due to Allen's ecological rule.[91]
Professor of anthropology at Trent University Ontario, Canada, Joseph K. So (1980) cited a study by J. T. Steegman (1965) that the so-called cold-adapted Mongoloid face has been shown in an experiment, using Japanese and European subjects, to not offer greater protection to frostbite.[92] In explaining Mongoloid cold-adaptiveness, So cites the work of W. L. Hylander (1977) where Hylander said that in the Eskimo (Inuit), for example, the reduction of the brow ridge and flatness of the face is due to internal structural configurations that are cold adapted in the sense that they produce a large vertical bite force necessary to chew frozen seal meat.[92]
.jpg)
Miquel Hernández of the Department of Animal Biology at the University of Barcelona said that the high and narrow nose of Eskimos (Inuit) and the Neanderthals is an adaptation to a cold and dry environment, since it contributes to warming and moisturizing the air and the "recovery of heat and moisture from expired air".[79]
A. T. Steegman of the Department of Anthropology at State University of New York investigated the assumption that Allen's rule caused the structural configuration of the Arctic Mongoloid face.[93] Steegman did an experiment that involved the survival of rats in the cold.[93] Steegman said that the rats with narrow nasal passages, broader faces, shorter tails and shorter legs survived the best in the cold.[93] Steegman paralleled his findings with the Arctic Mongoloids, particularly the Eskimo and Aleut, by claiming these Arctic Mongoloids have similar features in accordance with Allen's rule: a narrow nasal passage, relatively large heads, long to round heads, large jaws, relatively large bodies, and short limbs.[93]
Kenneth L. Beals of the Department of Anthropology at Oregon State University said that the indigenous people of the Americas have cephalic indexes that are an exception to Allen's rule, since the indigenous people of the hot climates of North and South America have cold-adapted, high cephalic indexes.[94] Beals said that these peoples have not yet evolved the appropriate cephalic index for their climate, being, comparatively, only recently descended from the cold-adapted Arctic Mongoloid.[94]
In 1950, Carleton S. Coon et al. said that Mongoloids have faces that are adapted to the extreme cold of subarctic and arctic conditions. Coon et al. said that Mongoloids have eye sockets that have been extended vertically to make room for the "heavy fat padding" that Mongoloids have around their eyeballs. Coon et al. said that Mongoloids have "reduced" brow ridges to decrease the size of the air spaces inside of their brow ridges known as the frontal sinuses which are "vulnerable" to the cold. Coon et al. said that Mongoloid facial features reduce the surface area of the nose by having nasal bones that are flat against the face and having enlarged cheekbones that project forward which effectively reduce the external projection of the nose.[59]
Physical features of "Asian" people
Willett Enos Rotzell professor of Botany and Zoology at the Hahnemann Medical College said the Asian race has skin color ranging from a yellowish tint to an olive shade, with black and coarse hair with a circular cross section, an absent or scanty beard, a brachycephalic skull, prominent cheek bones and a broad face. Rotzell said that the Asian race has its original home in Asia.[95]
Dennis C. Dirkmaat professor of paleoanthropology and archaeology at Mercyhurst University[96] said that Southeast Asian skulls can be distinguished from Asian and Native American skulls in that they are "smaller and less robust" with noses exhibiting a medium width without nasal overgrowth, and can "exhibit gracile features common to female skulls".[97]
Dr. Ann H. Ross, Co-Director of the Forensic Sciences Institute at North Carolina State University,[98] in a presentation on the concept of "race" (written in scare quotes) from the perspective of forensic anthropology, said individuals of "Asian ancestry" have an "intermediate profile", meaning the part of the maxilla is "moderate" compared to individuals of "African ancestry" who have a "projecting maxilla", and compared to individuals who are "White/Hispanic" who generally have a "straight profile" or "lack of prognathism". She qualified her statement about Hispanics by adding that their lack of prognathism would not hold true for Hispanic populations with "African admixture".[99]

Qing He et al. of the Obesity Research Center at Columbia University did a study on "fat distribution" of 358 prepubertal children and the study said that Asians had less gynoid fat than African Americans and more relative trunk fat than Caucasians, but less relative extremity fat than Caucasians.[100]
Douglas W. Deedrick, Unit Chief of the Trace Evidence Unit for the Federal Bureau of Investigation, said that hairs of "Mongoloid or Asian origin" are characterized as being straight and coarse with a circular cross section and a wider diameter than those of other "racial groups". He said that the cuticle is thicker than those of Negroid or Caucasian hairs while the medulla is "continuous and wide". He said that the pigment granules are smaller than the larger pigment granules of Negroid hair, and the pigment granules in the cortex are "generally larger" than those of Caucasian hair. Unlike the "evenly distributed" pigment granules of Caucasian hair, Asian hair frequently has clusters of pigment granules that form "patchy areas".[101]

Sang-Ki Jeong (Korean: 정상기) et al. of Chonnam University, Kwangju, Korea, using both Asian and Caucasian cadavers as well as four healthy young Korean men said that "Asian eyelids" whether "Asian single eyelids" or "Asian double eyelids" had more fat in them than in Caucasians.[103] Jeong et al. said that the cause of the "Asian single eyelid" was that "the orbital septum fuses to the levator aponeurosis at variable distances below the superior tarsal border; (2) preaponeurotic fat pad protusion and a thick subcutaneous fat layer prevent levator fibers from extending toward the skin near the superior tarsal border; and (3) the primary insertion of the levator aponeurosis into the orbicularis muscle and into the upper eyelid skin occurs closer to the eyelid margin in Asians."[103]

The average size of random melanosomes of "Asian skin" for Chinese individuals of Fitzpatrick phototype IV through V was measured to be 1.36 ± 0.15 μm2 x 10−2 which was between the higher value of 1.44 ± 0.67 μm2 x 10−2 measured for "African/American skin" of Fitzpatrick phototype VI and the lower value of 0.94 ± 0.48 μm2 x 10−2 measured for "Caucasian skin" of Fitzpatrick phototype II. The ratio of clustered to distributed melanosomes was 37.4% clustered vs. 62.6% distributed in Asian skin, 84.5%. clustered vs. 15.5% distributed in Caucasian skin and 11.1% clustered vs. 88.9% distributed in African/American skin.[104]
George Richard Scott, physical anthropologist at the University of Nevada, said that some East Asians (in particular, Han Chinese and some Japanese), as well as Native Americans, have a distinctive dental pattern known as Sinodonty, where, among other features, the upper first two incisors are not aligned with the other teeth, but are rotated a few degrees inward and are shovel-shaped.[105]
Both darker-skinned and lighter-skinned Asians have a thicker dermis than Caucasians of comparable skin pigment which may be the reason for a "substantially lower incidence of fine wrinkles" in Asians when compared to Caucasians, and this lower incidence of fine wrinkles may be the reason for the "myth" that Asian faces age slower than Caucasian faces.[106]
Asian people and black people have a thicker dermis than white people. The skin of Asian people and black people also has more sun protection than the skin of white people due to Asian people and black people having larger and more numerous melanosomes in their skin than white people. The thicker dermis and the more numerous melanosomes of larger size might be the reasons that Asian people and black people have a lower incidence facial wrinkles than white people.[107]
Turkic peoples


All the Turkic peoples native to Central Asia are of mixed Caucasoid and Mongoloid origin. Turkic people display a great variety of ethnic types.[108] They possess physical features ranging from Caucasoid to Northern Mongoloid. Mongoloid and Caucasoid facial structure is common among many Turkic groups, such as Chuvash people, Tatars, Kazakhs, Uzbeks, and Bashkirs. Historically, the racial classification of the Turkic peoples was sometimes given as "Turanid". Turanid racial type or "minor race", situated at the boundary of the distribution of the Mongoloid and Europid "great races".[109][110]
The Turkic people live in central, eastern, northern, and western Asia as well as parts of eastern Europe.[111] The term "Turkic" represents a broad ethnic group of peoples including existing societies such as Altai, Azerbaijanis, Balkars, Bashkirs, Chuvashes, Crimean Karaites, Gagauz, Karachays, Karakalpaks, Kazakhs, Khakas, Krymchaks, Kyrgyz people, Nogais, Qashqai, Tatars, Turkmens, Turkish people, Tuvans, Uyghurs, Uzbeks, and Yakuts and as well as past civilizations such as Dingling, Bulgars, Chuban, Göktürks, Oghuz Turks, Khazars, Khiljis, Kipchaks, Kumans, Karluks, Bahri Mamluks, Ottoman Turks, Seljuk Turks, Tiele, Timurids, Turgeshes, Yenisei Kirghiz, and possibly Huns, Tuoba, Wusun, and the Xiongnu.[111][112][113][114][115]
Mongolian spot
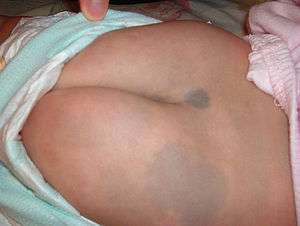
A Mongolian spot, also known as Mongolian blue spot, congenital dermal melanocytosis,[116] and dermal melanocytosis[116] is a benign, flat, congenital birthmark with wavy borders and irregular shape. In 1883 it was described and named after Mongolians by Erwin Bälz, a German anthropologist based in Japan.[117][118][119][120] It normally disappears three to five years after birth and almost always by puberty.[3] The most common color is blue, although they can be blue-gray, blue-black or deep brown.
The spot is prevalent among East, South, Southeast, North and Central Asian peoples, Indigenous Oceanians (chiefly Micronesians and Polynesians), Sub-Saharan Africans,[121] Amerindians,[122] non-European Latin Americans, Caribbeans of mixed-race descent, and Turkish people.[123][124][125][126] They occur in about 90-95% of Asian and 80-85% Native American infants.[125] Approximately 90% of Polynesians and Micronesians are born with Mongolian spots, as are about 46% of children in Latin America,[127] where they are associated with non-European descent. These spots also appear on 5-10% of babies of full Caucasian descent; Coria del Río in Spain has a high incidence due to the presence of descendants of the first Japanese official envoy to Spain in the early 17th century.[125][128] Black babies have Mongolian spots at a frequency of 96%.[129]
Genetic research
| Distances Between Races and Heterozygosity Based on Blood, Blood Groups, and Protein Enzymes | |||
|---|---|---|---|
| Genetic Criteria | Racial Composition | ||
| Caucasoid vs. Mongoloid | Mongoloid vs. Negroid | Negroid vs. Caucasoid | |
| Blood Group Loci | |||
| Number of Loci | 18 | 17 | 17 |
| Average Heterozygosity | 0.303 | 0.283 | 0.317 |
| Average Genetic Distance Between Races | 0.064 | 0.194 | 0.056 |
| Protein Enzyme Loci | |||
| Number of Loci | 38 | 40 | 41 |
| Average Heterozygosity | 0.210 | 0.202 | 0.217 |
| Average Genetic Distance Between Races | 0.025 | 0.065 | 0.067 |
| Source: Polednak (1989) who cited the data of Mourant et al. (1976), Tills et al. (1983a) & Roberts and De Stefano (1986)[130] | |||
| Genetic Distances and Effective Divergence Times Between The Three Major Races of Man (3) by Masatoshi Nei (Japanese: 根井正利), Professor of Biology at Pennsylvania State University[131] | ||||
|---|---|---|---|---|
| Comparison | Proteins (62 loci) |
Blood groups (23 loci) |
Total (85 loci) |
Effective divergence time (years) |
| Caucasoid/Mongoloid | 0.011 | 0.043 | 0.019 | 41,000 ± 15,000 |
| Caucasoid/Negroid | 0.030 | 0.038 | 0.032 | 113,000 ± 34,000 |
| Negroid/Mongoloid | 0.031 | 0.096 | 0.047 | 116,000 ± 34,000 |

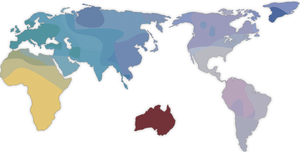

In 1994, geneticist Luigi Luca Cavalli-Sforza of Stanford University divided a principal coordinant map of 42 Asian populations into three groupings: Asian Caucasoids, Northeast and East Asian and Southeast Asian.[134] Along Southeast Asia, Cavalli-Sforza said that there is a separation between northern and southern Mongoloids.[134] To the West, Cavalli-Sforza said there is an approximate boundary between Caucasoids and Mongoloids from the Urals to the eastern part of India.[134] Along this boundary there has been hybridization, causing a Caucasoid-Mongoloid gradient.[134] More specifically, the ethnic groups Cavalli-Sforza said that were in the Northeast and East Asian cluster were the Koryak, Chukchi, Reindeer Chukchi, Nganasan Samoyed, Northern Tungus, Nentsy, N. Chinese, Tibetan, Bhutanese, Ainu, Mongol, Japanese and Korean.[134] Moving south, the ethnic groups Cavalli-Sforza said that were in the Southeast Asian cluster were the Indonesian, Malaysian, Taiwan aborigines, S. Chinese, Vietnamese, Muong, Thai, Filipino, Balinese and Nepalese.[134] Other studies also show that S. Chinese, Vietnamese and Tai peoples were found intermediate between the N. Chinese or other Northeast Asians and Southeast Asian.[135][136] "Reference populations".
Atsushi Tajima (田嶋敦) et al. of Graduate University for Advanced Studies, Hayama, Kanagawa, Japan, said that there is evidence for four separate populations, carrying distinct sets of non-recombining Y chromosome lineages, within the traditional Mongoloid category: North Asians, Han Chinese, Southeast Asians, and Japanese.[137]
Satoshi Horai (宝来聡) of the Japanese National Institute of Genetics, said that phylogenetic analysis indicated that there are two distinct groups of Mongoloids – one which early on diverged from Negroids and another that diverged from Caucasoids later.[138] Horai said that Mongoloid distribution corresponds to North and South America, Oceania, Southeast Asia, east Asia, and Siberia.[138]
A study conducted by the HUGO Pan-Asian SNP Consortium in 2009 used principal components analysis, which makes no prior population assumptions, on genetic data sampled from a large number of points across Asia. They said that East Asian and South-East Asian populations clustered together, and suggested a common origin for these populations. At the same time they observed a broad discontinuity between this cluster and South Asia, commenting most of the Indian populations showed evidence of shared ancestry with European populations. The study said that genetic ancestry is strongly correlated with linguistic affiliations as well as geography.[139]
Professor Lic. Edvin Santiago Trujillo said that antigen Dia is not present in blacks and Caucasians. Trujillo said that Dia is found in populations of Mongolic origin. Trujillo said that Dia is present in Chinese (2 — 5%), Polish (0.25 — 0.91%), Japanese (8 — 12%), Mexican Americans (8.2 — 14.7%) and American Indian tribes (7 — 54%). Trujillo said that when Dia is found in Caucasians, there is a presumption that they have a Mongoloid ancestor.[140]
| "Estimates of the Number of Nucleotide Differences per Site Both Among (dxy) and within (dx or dy) Each of the Three Races, and Net Nucleotide Differences (d) among the Races" made by Satoshi Horai (Japanese:宝来聡) of the Department of Human Genetics, National Institute of Genetics, Mishima, Shizouka, Japan.[138] | ||||
|---|---|---|---|---|
| Caucasoid (N=20) |
Mongoloid (N=71) |
Negroid (N=10) | ||
| Caucasoid | 0.0094 | 0.0012 | 0.0028 | |
| Mongoloid | 0.0128 | 0.0137 | 0.0015 | |
| Negroid | 0.0194 | 0.0203 | 0.0238 | |
DNA studies of "Asian" populations
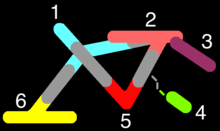
Peter A. Underhill, Department of Genetics, Stanford University and Toomas Kivisild of the Leverhulme Center of Human Evolutionary Studies, University of Cambridge[141]
1. Europe 2. East Asia 3. Americas 4. Australia and Oceania | 5. South Asia 6. East Africa transition |
Scott W. Ballinger et al. of the Department of Biochemistry at Emory University said "Asian mtDNA lineages" originated in Southern China with the "Southern Mongoloid".[142]
Hiroki Oota (太田博樹) et al. of the Max Planck Institute for Evolutionary Anthropology, Germany, said that "Asian populations" have high mtDNA variation with Vietnamese having the highest mtDNA diversity, but, overall, the genetic distance between "Asian populations" is small.[143]
Melissa L. Cann et al. of the Department of Biochemistry at the University of California, Berkeley, said that early Asians did not mix with "Asian Homo and that the features of "ancient Asian forms" indicate that "Asian erectus" was not ancestral to "Homo sapiens".[144] Since modern-day "Asians" do not show the amount of mtDNA divergence expected had they mixed with Homo erectus, Cann believes the expanding Homo sapiens from Africa replaced the Asian Homo erectus.[144]
Douglas C. Wallace of the Department of Biochemistry at Emory University said that the mtDNA of the indigenous peoples of the Americas is "clearly Asian in character", but the few founding females carried "rare Asian mtDNAs", causing a different frequency of mtDNA and a "dramatic founder effect".[145] The Austro-Asiatic groups of India are proto Asiatic groups, like the Munda people.
Shama Barnabas, B. Joshi and C.G. Suresh of the Division of Biochemical Sciences, National Chemical Laboratory, Pune, India, said that evidence for the original people of India who they refer to as the "proto-Asiatic element" spreading into Southeast Asia to become Southeast Asians is shown by the mtDNA affinities between Indians and East Asians and Southeast Asians in DdeI 10394 site along with the associated Asian-specific AluI 10397 site.[146]
| Data from Simon Gravel et al. (2010)[147] | ||||||
|---|---|---|---|---|---|---|
| Divergence Event | ||||||
| Data from NIEHS | Low-Coverage + Exons | Exons | ||||
| European and Asian Divergence Event | 21,200 years ago | 23,000 years ago | 28,000 years ago | |||
| Eurasian and African Divergence Event | 140,000 years ago | 51,000 years ago | 98,000 years ago | |||
| African and Ancestral Divergence Event | 220,000 years ago | 148,000 years ago | 316,000 years ago | |||
| The African population used in this study was Yoruba people in Ibadan, Nigeria. The Asian populations used in this study were Han Chinese people in Beijing and Japanese people in Tokyo. The European population used in this study was European-origin people in Utah. The acronym "NIEHS" stands for the National Institute of Environmental Health Sciences. The column header "Low-Coverage + Exons" means a combination of "whole-genome, low-coverage sequencing" and the "exon pilot data". | ||||||
| Genetic diversity within/between continental populations by Hiroki Oota et al. of the Max Planck Institute for Evolutionary Anthropology, Germany[143] | |||
|---|---|---|---|
| Number of Populations | Within Populations average mean pairwise differences | Between population average Fst | |
| Africa | 15 | 7.99 ± 2.72 | 0.201 |
| Europe | 12 | 4.63 ± 0.94 | 0.066 |
| Asia | 12 | 7.12 ± 0.91 | 0.033 |
| Eurasia | 27 | 5.95 ± 1.51 | 0.086 |
| mtDNA divergence within and between 5 human populations by Melissa L. Cann et al. of the Department of Biochemistry at the University of California, Berkeley[144] | |||||
|---|---|---|---|---|---|
| Population | 1 | 2 | 3 | 4 | 5 |
| 1. African | 0.47 | 0.04 | 0.04 | 0.05 | 0.06 |
| 2. Asian | 0.45 | 0.35 | 0.01 | 0.02 | 0.04 |
| 3. Australian | 0.40 | 0.31 | 0.25 | 0.03 | 0.04 |
| 4. Caucasian | 0.40 | 0.31 | 0.27 | 0.23 | 0.05 |
| 5. New Guinean | 0.42 | 0.34 | 0.29 | 0.29 | 0.25 |
| The divergence is calculated by a way developed by Masatoshi Nei. The values of the mean pairwise divergence between individuals within populations (δx) appear on the diagonal. The values below the diagonal (δxy) are the mean pairwise divergences between individuals belonging to two different populations, X and Y. The values above the diagonal (δ) are interpopulation divergences that corrected for variation within those populations with the equation δ = δxy – 0.5(δx + δy). | |||||
Criticism
Dr. George W. Gill, professor of anthropology at the University of Wyoming and Dennis O'Neil professor of anthropology at Palomar College, said that "Mongoloid" concept originated with a now disputed typological method of racial classification.[148][149] All the -oid racial terms (e.g. Mongoloid, Caucasoid, Negroid, etc.) are now often controversial in both technical and non-technical contexts and may sometimes give offense no matter how they are used.[150]
According to Ward O. Conner who wrote a book about John Langdon Down, since people with Down syndrome may have epicanthic folds, Down syndrome was widely called "Mongol" or "Mongoloid Idiocy".[151] In reference to this, the term grew in usage as a pejorative. A shortened version of the term, mong or mongol, is also used in the United Kingdom.[152] Chong Yah Lim, professor of economics at Nanyang Technological University in Singapore said that he did not like the term "Mongoloid" because it has "come to mean demented physical and mental developments". Lim said that he thought the term "East Asian race" would be a more "appropriately neutral, modern term".[153]
Gallery
-

East Asian types in a book from 1914.
-

Natives of North America in a book from 1914.
-

Natives of South America in a book from 1914.
-

Dayak man from Borneo, Southeast Asia in 1900s.
-

Young Māori woman with traditional tattoos from New Zealand (Aotearoa).
-

Sea Sami man from Northern Norway.^
-

Kalmyks, 19th century.

See also
| Look up mongoloid in Wiktionary, the free dictionary. |
| Wikimedia Commons has media related to Race. |
- Afro-Asian (mixed ancestry)
- Asian people
- Craniofacial anthropometry
- Eurasian (mixed ancestry)
- Mestizo (mixed ancestry)
- Mongols
References
- ↑ Mongoloid. (2012). Dictionary.com. Retrieved September 3, 2012, from link.
- ↑ For a contrast with the "Europoid" or Caucasian race see footnote #4 of page 58-59 in Beckwith, Christopher. (2009). Empires of the Silk Road: a History of Central Eurasia from the Bronze Age to the Present. Princeton and Oxford: Princeton University Press. ISBN 978-0-691-13589-2.
- 1 2 Mongolian Spot DrGreen.com
- ↑ Mongolian Spot - English information of Mongolian spot, written by Hironao NUMABE, M.D., Tokyo Medical University.
- 1 2 Montagu, A. (1951). An introduction to physical anthropology: A revised second edition. Charles C. Thomas Publisher: Springfield, Illinois, USA.
- ↑ Roth, PL; Bevier, CA; Bobko, P; Switzer, FS, III; Tyler, P (2001). "Ethnic group differences in cognitive ability in employment and educational settings: A metaanalysis". Personnel Psychology. 54: 297–330. doi:10.1111/j.1744-6570.2001.tb00094.x.
- ↑ Ho, KC; Roessmann, U; Straumfjord, JV; Monroe, G (December 1980). "Analysis of brain weight. I. Adult brain weight in relation to sex, race, and age.". Archives of Pathology & Laboratory Medicine. 104 (12): 635–9. PMID 6893659.
- ↑
- Lieberman, Leonard (2001). "How "Caucasoids" got such big crania and why they shrank: from Morton to Rushton."". Current Anthropology. 42 (1): 69–95. doi:10.1086/318434. PMID 14992214.
- 1 2 Smay, Diana; Armelagos, George. "Galileo Wept: A Critical Assessment of the Use of Race in Forensic Anthropology" (PDF). Emory University.
- 1 2 Lieberman, Leonard. "Out of Our Skulls: Caucasoid, Mongoloid, Negroid?". Anthropology News. 38: 56. doi:10.1111/an.1997.38.9.56.
- 1 2 Templeton, Alan R. "Human Races: A Genetic and Evolutionary Perspective" (PDF). Washington University. Realfuture.org.
- 1 2 Keevak, Michael. "Becoming Yellow: A Short History of Racial Thinking". Princeton: Princeton University Press, 2011. ISBN 978-0-691-14031-5.
- ↑ Diamond, JM (2000). "Taiwan's gift to the world". Nature. 403 (6771): 709–710. doi:10.1038/35001685. PMID 10693781.
- ↑ Gunn, Bee; Luc Baudouin; Kenneth M. Olsen (2011). "Independent Origins of Cultivated Coconut (Cocos nucifera L.) in the Old World Tropics". PLoS ONE. 6 (6): e21143. doi:10.1371/journal.pone.0021143. PMC 3120816
 . PMID 21731660.
. PMID 21731660. - ↑ Van Tilburg, Jo Anne. 1994. Easter Island: Archaeology, Ecology and Culture. Washington D.C.: Smithsonian Institution Press
- ↑ Langdon, Robert. The Bamboo Raft as a Key to the Introduction of the Sweet Potato in Prehistoric Polynesia, The Journal of Pacific History', Vol. 36, No. 1, 2001
- 1 2 L.E. Beckman, K. Sjoberg, S. Eriksson and L. Beckman (2001). "Haemochromatosis gene mutations in Finns, Swedes and Swedish Saamis". Human Heredity. 52 (2): 110–112. doi:10.1159/000053362. PMID 11474212.
- ↑ Meyers Konversations-Lexikon, 4th edition, 1885-90.
- ↑ Warren, D.M. (1856). A System of Physical Geography. Philadelphia: H. Cowperthwait & Co. pp. 77.
- ↑ Winchell, A. (1881). Preadamites; or A Demonstration of the Existance of Men Before Adam; (3rd ed.). Chicago: S.C. Griggs and Company; London: Trubner & Co. pp. 57.
- ↑ Berg, M. & Wendt, S. (2014). Racism in the Modern World: Historical Perspectives on Cultural Transfer and Adaptation. Berghahn Books. pp. 239. ISBN 978-1-78238-086-3
- ↑ Robbins, C.R. (1994). Chemical and Physical Behavior of Human Hair (3rd ed.). New York: Springer Science. pp. 328. ISBN 978-1-4757-3898-8
- ↑ Painter, Nell Irvin (2003). "Why White People are Called Caucasian?" (PDF). Yale University. Retrieved September 27, 2007. Keevak. Becoming Yellow, pp. 74–77
- ↑ Blumenbach, Johann. "The Anthropological Treatise of Johann Friedrich Blumenbach". London: Longman Green, 1865.
- 1 2 Deniker, Joseph. The Races of Man: An Outline of Anthropology and Ethnography C. Scribner's Sons: New York, 1900. ISBN 0-8369-5932-9
- ↑ [The End of Racism by Dinesh D'Souza, pg 124]
- ↑ Gobineau, Arthur (1915). The Inequality of Human Races. Putnam. ISBN 0-86527-430-4. Retrieved 2007-10-18.
- ↑ DiPiero, Thomas. White Men Aren't gid/s work Duke University Press, 2002. ISBN 0-8223-2961-1
- ↑ "Huxley, Thomas, On the Geographical Distribution of the Chief Modifications of Mankind. 1870. August 14, 2006". Aleph0.clarku.edu. Retrieved 2013-12-15.
- ↑ James Dallas, "On the Primary Divisions and Geographical Distributions of Mankind", 1886 Royal Anthropological Institute of Great Britain and Ireland, p.304-30. James describes this as "equivalent to Professor Huxley's Mongoloid division" and as encompassing "Mongols and American Indians"
- 1 2 Boas, F. (1940). Race, language, and culture. New York: Macmillan.
- ↑ Anemone, R. L. (1996). Obituary: Elizabeth Smithgall Watts (1941-1994). In American Journal of Physical Anthropology. (99)221-222. link
- ↑ Watts, E.S. (1981). "The Biological Race Concept and Diseases of Modern Man." In Biocultural Aspects of Disease. New York: Academic Press.
- 1 2 Futuyma, Douglas A. Evolutionary Biology. Massachusetts: Sinauer Associates, 1983. p. 520
- ↑ "University Catalog". California State University, Chico. 2003. Retrieved September 28, 2007.
- ↑ Lederer Roger J. Ecology and Field Biology. Cummings Publishing Company: California, 1984. ISBN 0-8053-5718-1 p.129
- 1 2 3 Lahr M. M. (1995). "Patterns of modern human diversification: Implications for Amerindian origins". American Journal of Physical Anthropology. 38: 163–198. doi:10.1002/ajpa.1330380609.
- ↑ Niskanen, M. (2002). The Origin of the Baltic-Finns from the Physical Anthropological Point of View. Mankind Quarterly Volume XLIII Number 2, Winter. p. 125
- ↑ http://arxiv.org/ftp/arxiv/papers/1312/1312.6639.pdf
- ↑ Bellwood, Peter. Pre-History of the Indo-malaysian Archipelago. Australian National University:1985. ISBN 978-1-921313-11-0
- 1 2 3 4 5 6 7 Takeru Akazawa and Emóke J.E. Sathmåry. Prehistoric Mongoloid dispersals. New York, Oxford University Press, 1996.
- ↑ Tarling, N. (1992). The Cambridge History of Southeast Asia: From Early Times to c. 1800 (Vol. 1). United Kingdom: Cambridge University Press. 74-77.
- ↑ Huxley, Thomas. Collected Essays of Thomas Huxley: Man's Place in Nature and Other Kessinger Publishing: Montana, 2005. ISBN 1-4179-7462-1
- ↑ Sangvichien, S. (1964). A Preliminary Report on Non-Metrical Characteristics of Neolithic Skeletons Found at Ban Kao, Kanchanaburi. page 8. Retrieved November 11, 2016, from link
- ↑ Coon, Carleton S. The Races of Europe. Greenwood: USA, 1972 ISBN 0-8371-6328-5 p.2
- 1 2 3 Coon, Carleton S. The Origin of the Races. Knopf: Michigan, 1962. ISBN 0-394-30142-0
- ↑ Huxley, T. H. On the Geographical Distribution of the Chief Modifications of Mankind (1870) Journal of the Ethnological Society of London. Huxley indicates that he has omitted certain areas with complex ethnic compositions that do not fit into his racial paradigm, including much of the Indian subcontinent and Horn of Africa. (Huxley, Thomas (1873). Critiques and Addresses by Thomas Henry Huxley, LL.D., F.R.S. Macmillan and Company. p. 153.) By the late nineteenth century, his Xanthochroi group had been redefined as the Nordic race, whereas his Melanochroi became the Mediterranean race. As such, Huxley's Melanochroi eventually also comprised various other dark Caucasoid populations, including the Hamites and Moors. (Gregory, John Walter (1931). Race as a Political Factor. Watts & Company. p. 19. Retrieved 8 May 2016.)
- ↑ Bhasin, M.K. (2006). "Genetics of Caste and Tribes of India: Indian Population Milieu" (PDF). Int J Hum Genet. Kamla Raj. 6 (3): 244. Retrieved 2007-10-22.
- ↑ Milford Wolpoff & Rachel Caspari (1998). Race and Human Evolution: A Fatal Attraction. Westview Press. ISBN 0-8133-3546-9.
- ↑ Peter Brown (1999). "The First Modern East Asians? another Look at Upper Cave 101, Liujiang, and Minatogawa" (PDF). K. Omoto (ed.) Interdisciplinary Perspectives on the Origins of the Japanese, International Research Center for Japanese Studies: Kyoto. Department of Anthropology and Paleoanthropology, University of New England. pp. 105–130. Retrieved 2007-09-23.
- ↑ Yali Xue,*,†,‡ Tatiana Zerjal,*,‡ Weidong Bao,‡,§ Suling Zhu,‡,§ Qunfang Shu,§ Jiujin Xu,§ Ruofu Du,§ Songbin Fu,† Pu Li,† Matthew E. Hurles,* Huanming Yang** and Chris Tyler-Smith*,‡,1 (2006). "Male Demography in East Asia: A North–South Contrast in Human Population Expansion Times" (PDF). Genetics Society of America. doi:10.1534/genetics.105.054270. Retrieved 2007-09-29.
- ↑ Posnansky, A. (1917). Signos Mongoloides en Algunos Tipos Étnicos del Altiplano Andino. In Proceedings of the Second Pan American Scientific Congress:(section I) Anthropology. WH Holmes, chairman (Vol. 1, p. 112). US Government Printing Office.
- ↑ Laughlin, W.S. (1963). Eskimos and Aleuts: Their Origins and Evolution. In Science, (142)3593. Page 639. Retrieved December 3, 2016, from link.
- ↑ Kolas, S. et al. (1953). The occurrence of torus palatinus and torus mandibularis in 2,478 dental patients [Abstract]. In Oral Surgery, Oral Medicine, Oral Pathology, and Oral Radiology, (6)9, pp. 1134-1141. Retrieved December 3, 2016, from link.
- ↑ Wu, R. & Olsen, J.W. (2009). Paleoanthropology and Paleolithic Archaeology in the People's Republic of China. USA: Left Coast Press, Inc. Page 116.
- ↑ Conceição, L. et al. (2015). Forensic Dentistry: An Overview of the Human Identification’s Techniques of this Dental Specialty. In Journal of Forensic Research, 6(1). Page 2. Retrieved November 30, 2016, from link.
- ↑ Thali, M.J., Viner, M.D. & Brogdon, B.G. (2011). Brogdon's Forensic Radiology, Second Edition. USA: CRC Press. Page 101.
- ↑ Ubelaker, D.H. & Scammell, H. (1992). Bones: A Forensic Detective's Casebook. New York: M. Evans and Company, Inc. Page 93.
- 1 2 Dahlberg, A.A. & Graber, T.M. (1977). Orofacial Growth and Development. USA & Canada: Mouton Publishers. Pages 132, 147 & 148.
- ↑ Clark, G.A. & Willermet, C.M. (1997). Conceptual Issues in Modern Human Origins Research. New York: Aldine de Gruyter. page 195.
- ↑ Gill, George W. 1998. "Craniofacial Criteria in the Skeletal Attribution of Race. " In Forensic Osteology: Advances in the Identification of Human Remains. (2nd edition) Reichs, Kathleen l(ed.), pp. 293–315.
- ↑ Diana Smay, George Armelagos (2000). "Galileo wept: A critical assessment of the use of race of forensic anthropolopy" (PDF). Transforming Anthropology. 9 (2): 22–24. Retrieved 13 July 2016.
- ↑ Adhikari, K., Fuentes-Guajardo, M., Quinto-Sánchez, M., Mendoza-Revilla, J., Chacón-Duque, J. C., Acuña-Alonzo, V., Gómez-Valdés, J. (2016). "A genome-wide association scan implicates DCHS2, RUNX2, GLI3, PAX1 and EDAR in human facial variation". Nature communications. 7. Retrieved 12 November 2016.
- 1 2 Blumenfield J (2000). "Racial Identification in the Skull and Teeth". Totem: The University of Western Ontario Journal of Anthropology. 8 (1): 21–33.
- ↑ Yaacob, H., Narnbiar, P. & Naidu, M.D.K. (1996). Racial characteristics of human teeth with special emphasis on the Mongoloid dentition. Malaysian Journal of Pathology, 18(1), 5.
- 1 2 Stefan, V.H. & Gill, G.W. (2016). Skeletal Biology of the Ancient Rapanui (Easter Islanders). United Kingdom: Cambridge University Press. 275-277.
- 1 2 Wilkinson, C. (2004). Forensic Facial Reconstruction. Cambridge University Press. ISBN 0-521-82003-0
- ↑ Ong RG, Stevenson MR (January 1999). "Evaluation of bone density in the mandibles of young Australian adults of Mongoloid and Caucasoid descent". Dentomaxillofacial Radiology. 28 (1): 20–5. doi:10.1038/sj.dmfr.4600399. PMID 10202474.
- ↑ Sullivan, L.R., Gifford, E.W. & McKern, W.C. (1921). A Contribution to Samoan Somatology. Bishop Museum Press: Hawaii. Pages 94, 97 & 98.
- 1 2 Wu, Ju-kang (1959). "Human Fossils Found in Liukiang, Kwangsi, China" (PDF). Paleovertebrata et Paleoanthropologia. 1 (3): 97–104.
- ↑ Yuzurihaa Shunsuke; Matsuo Kiyoshi; Kushimaa Hideo (2000). "An anatomical structure which results in puffiness of the upper eyelid and a narrow palpebral fissure in the Mongoloid eye.". British Journal of Plastic Surgery. 53 (6): 466–472. doi:10.1054/bjps.2000.3387.
- 1 2 Schurr, Theodore G. (2011). Mapping Mongolia: Situating Mongolia in the World from Geologic Time to the Present. University of Pennsylvania Museum of Archaeology and Anthropology. Pennsylvania. ISBN 1-934536-18-0
- ↑ Dawber R.P.R. (1997). Diseases of the head and scalp (3rd ed.). Virginia:Blackwell Science Ltd.
- ↑ Trotter M (1938). "A review of the classifications of hair". American Journal of Physical Anthropology. 24: 105–126. doi:10.1002/ajpa.1330240131.
- ↑ Duggins O. H.; Trotter M.; Coon C. S. (1959). "Hair from a Kadar woman of India". American Journal of Physical Anthropology. 17: 95–98. doi:10.1002/ajpa.1330170203.
- ↑ Hrdy D (1973). "Quantitative hair form variation in seven populations". American Journal of Physical Anthropology. 39: 7–17. doi:10.1002/ajpa.1330390103.
- 1 2 Haydenblit R (1996). "Dental variation among four prehispanic Mexican populations". American Journal of Physical Anthropology. 100: 225–246. doi:10.1002/(SICI)1096-8644(199606)100:2<225::AID-AJPA5>3.0.CO;2-W.
- ↑ Robert B. Pickering, David Bachman. (2009). The Use of Forensic Anthropology (2nd ed.). CRC Press. ISBN 9781420068771. pp. 83
- 1 2 Hernández M.; Fox C. L.; Garcia-Moro C. (1997). "Fueguian cranial morphology: The adaptation to a cold, harsh environment". American Journal of Physical Anthropology. 103: 103–117. doi:10.1002/(SICI)1096-8644(199705)103:1<103::AID-AJPA7>3.0.CO;2-X.
- ↑ Coon, C.S. (1939). The Races of Europe. USA: The Macmillan Company.
- ↑ Berardesca, E., Lévêque, J. & Maibach H.I. (2007). Ethnic Skin and Hair (Dermatology: Clinical & Basic Science). USA: Informa Healthcare, Inc. 82, 88 & 89.
- ↑ Franz Boas. (1905). Anthropometry of Central California. Harvard University.
- ↑ Hudson, Mark J. (1999). Ruins of identity: ethnogenesis in the Japanese Islands
- ↑ Aleš Hrdlička. 1906. Contribution to the Physical Anthropology of California. Berkeley University Press.
- ↑ Oppenheimer, Stephen. The Real Eve. Published by Carroll & Graf Publishers, 2003 ISBN 0-7867-1192-2
- 1 2 Storm, P. The evolutionary significance of the Wajak skulls. — Scripta Geol., 110: 1-247, figs. 1-30, tabs. 1-121, Leiden, September 1995.
- ↑ Ronald Elmslie and Susan Nance, 'Abbie, Andrew Arthur (1905–1976)', Australian Dictionary of Biography, National Centre of Biography, Australian National University, link, published first in hardcopy 1993, accessed online 18 November 2014.
- ↑ Abbie A.A. (1964). "THE FACTOR TIMING IN EMERGENCE DISTINCTIVELY HUMAN CHARACTERS". Papers and proceedings of the Royal Society of Tasmania. 98: 63–71.
- ↑ Nicholas, Wade. Before the Dawn. Published by Penguin Publishing, 2006 ISBN 978-1594200793
- ↑ "秋田大学: Comparison between Triceps and Subscapular Skinfold Thickness of Japanese Children with respect to Environmental Effects". Air.lib.akita-u.ac.jp. Retrieved 2013-12-15.
- ↑ Takasaki Yuji; Loy Steven F.; Juergens Hans W. (2003). "Ethnic Differences in the Relationship between Bioelectrical Impedance and Body Size". Journal of Physiological Anthropology and Applied Human Science. 22: 233–235. doi:10.2114/jpa.22.233.
- 1 2 Joseph K. So. "Human Biological Adaptation to Arctic and Subarctic Zones". Annual Review of Anthropology; Vol. 9, (1980), pp. 63–82
- 1 2 3 4 Steegmann A. T.; Platner W. S. (1968). "Experimental cold modification of cranio-facial morphology". American Journal of Physical Anthropology. 28: 17–30. doi:10.1002/ajpa.1330280111.
- 1 2 Beals K. L. (1972). "Head form and climatic stress". American Journal of Physical Anthropology. 37: 85–92. doi:10.1002/ajpa.1330370111.
- ↑ Willett Enos Rotzell. (1905). Man: an introduction to anthropology. Philadelphia.
- ↑ "Dennis C. Dirkmaat, Ph.D., D.A.B.F.A.". Mai.mercyhurst.edu. Retrieved 20 January 2015.
- ↑ Dirkmaat, D. (2012). A companion to forensic anthropology. Wiley-Blackwell Publishing: USA. pp. 300 ISBN 978-1-4051-9123-4
- ↑ Ann H Ross. Research Gate Retrieved March 8, 2014, from Research Gate
- ↑ Ross, A.H. (2011). Seminar Session 1 The Concept of "Race": A Forensic Anthropological Perspective on Human Variation. Advances in Forensic Anthropology Retrieved March 8, 2014, from You Tube Video
- ↑ Horlick M; Thornton J; Wang J; Pierson R.N.; Heshka S; Gallagher D (2002). "Sex and race differences in fat distribution among Asian, African-American and Caucasian prepubertal children". Journal of Clinical Endocrinology and Metabolism. 87 (5): 2164–2170. doi:10.1210/jc.87.5.2164.
- ↑ Deedrick D. W. (2000). "Hairs, fibers, crime, and evidence". Forensic science communications. 2: 3.
- ↑ Bellwood, Peter (1985). Prehistory of the Indo-Malaysian Archipelago. Australian National University. p. 92. ISBN 978-1-921313-11-0.
- 1 2 Jeong Sangki; Lemke Bradley N.; Dortzbach Richard K.; Geol Park Yeoung; Keun Kang Heoung (1999). "The Asian Upper Eyelid: An Anatomical Study With Comparison to the Caucasian Eyelid". Arch Ophthalmol. 117: 907–912. doi:10.1001/archopht.117.7.907. PMID 10408455.
- ↑ Thong H. Y.; Jee S. H.; Sun C. C.; Biossy R. E. (2003). "The patterns of melanosome distribution in keratinocytes of human skin as one determining factor of skin colour". British Journal of Dermatology. 149 (3): 498–505. doi:10.1046/j.1365-2133.2003.05473.x. PMID 14510981.
- ↑ George Richard Scott; Christy G. Turner (1997). The Anthropology of Modern Human Teeth: Dental Morphology and Its Variation. Cambridge University Press.
- ↑ McCurdy, J.A., Jr., & Lam, S.M. (2005). Cosmetic Surgery of the Asian Face (2nd ed.). China: Thieme Medical Publishers, Inc. pp. 4. TMP ISBN 1-58890-218-8 GTV ISBN 3 13 747602 X
- ↑ Goldman, M.P. et al. (2013). Lasers and Energy Devices for the Skin (2nd ed.). Taylor & Francis Group. pp. 293. ISBN 978-1-84184-934-8
- ↑ Turkic people, Encyclopædia Britannica, Online Edition, 2008
- ↑ Racial and cultural minorities: an analysis of prejudice and discrimination, Environment, development, and public policy, George Eaton Simpson, John Milton Yinger, Springer, 1985, ISBN 0-306-41777-4, p.32.
- ↑ American anthropologist, American Anthropological Association, Anthropological Society of Washington (Washington, D.C,), 1984 v. 86, nos. 3-4, p. 741.
- 1 2 Turkic people, Encyclopædia Britannica, Online Academic Edition, 2010
- ↑ Pritsak O. & Golb. N: Khazarian Hebrew Documents of the Tenth Century, Ithaca: Cornell Univ. Press, 1982.
- ↑ "Timur", The Columbia Encyclopedia, Sixth Edition, 2001–05, Columbia University Press.
- ↑ Encyclopædia Britannica article: Consolidation & expansion of the Indo-Timurids, Online Edition, 2007.
- ↑ Walton, ,Linda (2013). World History: Journeys from Past to Present. p. 210.
- 1 2 Rapini, Ronald P.; Bolognia, Jean L.; Jorizzo, Joseph L. (2007). Dermatology: 2-Volume Set. St. Louis: Mosby. p. 1720. ISBN 1-4160-2999-0.
- ↑ Die koerperlichen Eigenschaften der Japaner.(1885) Baelz.E. Mittheil.d.deusch Gesell.f.Natur-u-Voelkerheilkunde Ostasiens. Bd.4.H.32
- ↑ Circumscribed dermal melanosis (Mongolian spot)(1981) Kikuchi I, Inoue S. in "Biology and Diseases of Dermal Pigmentation", University of Tokyo Press , p83
- ↑ Bernard Cohen (1993). Atlas of pediatric dermatology. Wolfe. pp. 6–17. ISBN 1563750198. Retrieved May 17, 2014.
- ↑ JAMA: The Journal of the American Medical Association, Volume 51. American Medical Association. American Medical Association. 1908. p. 2262. Retrieved May 17, 2014.
- ↑ Kevin C. Stuart (1997). Mongols in Western/American consciousness (illustrated ed.). Edwin Mellen Press. p. 95. ISBN 0773484434. Retrieved May 17, 2014.
- ↑ Miller (1999). Nursing Care of Older Adults: Theory and Practice (3, illustrated ed.). Lippincott Williams & Wilkins. p. 90. ISBN 0781720761. Retrieved May 17, 2014.
- ↑ Mongolian blue spots - Health care guide discussing the Mongolian blue spot.
- ↑ "Frequency and characteristics of Mongolian spots among Turkish children in Aegean region". July 2006.
- 1 2 3 "About Mongolian Spot". tokyo-med.ac.jp. Retrieved 1 October 2015.
- ↑ Transcultural Medicine: Dealing with patients from different cultures
- ↑ Epidemiology of Mongolian spot on MedScape
- ↑ "Spain's Japon clan has reunion to trace its 17th century roots - The Japan Times". The Japan Times. Retrieved 1 October 2015.
- ↑ N Silverberg (2012). Atlas of Pediatric Cutaneous Biodiversity: Comparative Dermatologic Atlas of Pediatric Skin of All Colors. Springer Science & Business Media. p. 34. ISBN 1461435641. Archived from the original on 2012. Retrieved May 17, 2014.
- ↑ Polednak, A.P. (1989). Racial & Ethnic Differences in Disease. New York and Oxford: Oxford University Press. 36.
- ↑ Nei, M. (1985). Human Evolution at the Molecular Level. Population Genetics and Molecular Evolution. Japan Sci. Soc. Press, Tokyo. pp. 44–64.
- 1 2 "斎藤成也 Naruya, S. Kyushu Museum. 2002. February 2, 2007". Museum.kyushu-u.ac.jp. Retrieved 2013-12-15.
- 1 2 Roberts, D.F., Fujiki, N. and Torizuka, N. (1992). Isolation, Migration and Health. 33rd Symposium Volume of the Society for Study of Human Biology.
- 1 2 3 4 5 6 7 Cavalli-Sforza, L.L., Menozzi, P. & Piazza, A. (1994). The History and Geography of Human Genes. New Jersey: Princeton University Press.
- ↑ Table from "Genetic evidence supports demic diffusion of Han culture". Nature (journal, 16 September 2004 issue)
- ↑ Table from " A spatial analysis of genetic structure of human populations in China reveals distinct difference between maternal and paternal lineages". European Journal of Human Genetics (journal, 23 January 2008 issue)
- ↑ Tajima A; PAN I.-H; Fucharoen G; Fucharoen S; Matsuo M; Tokunaga K; Juji T; Hayami M; Omoto K; Horai S (2002). "Three major lineages of Asian Y chromosomes: implications for the peopling of east and southeast Asia". Human Genetics. 110 (1): 80–88. doi:10.1007/s00439-001-0651-9.
- 1 2 3 Horai Satoshi; Hayasaka Kenji (1990). "Intraspecific Nucleotide Sequence Differences in the Major Noncoding Region of Human Mitochondrial DNA". Am. J. Hum. Genet. 46: 828–842.
- ↑ Mapping Human Genetic Diversity in Asia, The HUGO Pan-Asian SNP Consortium, 2009
- ↑ Grupo Cooperativo Iberoamericano de Medicina Transfusional.link
- ↑ Underhill Peter A.; Kivisild Toomas (2007). "Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations". Annual Review of Genetics. 41: 539–564. doi:10.1146/annurev.genet.41.110306.130407. PMID 18076332.
- ↑ Ballinger S.W. (1992). "Southeast Asian Mitochondrial DNA Analysis Reveals Genetic Continuity of Ancient Mongoloid Migrations". Genetics. 130: 139–152.
- 1 2 Hiroki Oota; Kitano Takashi; Jin Feng; Yuasa Isao; Wang Li; Ueda Shintaroh; Saitou Naruya; Stoneking Mark (2002). "Extreme mtDNA Homogeneity in Continental Asian Populations". American Journal of Physical Anthropology. 118: 146–153. doi:10.1002/ajpa.10056.
- 1 2 3 Rebecca L. Cann, Mark Stoneking, and Allan C. Wilson. Mitochondrial. (1987). DNA and Human Evolution. Letters to Nature
- ↑ Wallace D.C.; Garrison K; Knower W.C. (1985). "Dramatic Founder Effects in Amerindian Mitochondrial DNAs". American Journal of Physical Anthropology. 68: 149–155. doi:10.1002/ajpa.1330680202.
- ↑ Barnabas S.; Joshi B.; Suresh C. G. "Indian-Asian Relationship: mtDNA Reveals More". Naturwissenschaften. 87 (4): 180–183. doi:10.1007/s001140050699.
- ↑ Gravel S.; et al. (2010). "Demographic history and rare allele sharing among human populations" (PDF). Proceedings of the National Academy of Sciences of the United States of America. 108 (29): 11983.
- ↑ O'Neil, Dennis (May 13, 2007). "Biological Anthropology Terms". Palomar College. Check date values in:
|year= / |date= mismatch(help) - ↑ Gill, George W. "Does Race Exist? A proponent's perspective". Pbs.org.
- ↑ "Bartleby.com: Great Books Online -- Quotes, Poems, Novels, Classics and hundreds more". Bartleby.com. Retrieved 20 January 2015.
- ↑ Ward, Connor O. John Langdon (2006). "Down the man and the message". Down-syndrome.info.
- ↑ Clark, Nicola (October 19, 2011). "Ricky Gervais, please stop using the word 'mong'". London: The Guardian. Retrieved 26 May 2012.
- ↑ Chong Yah Lim. Southeast Asia: The Long Road Ahead. World Scientific, 2004 P. 3.