Plant cell

Plant cells are eukaryotic cells that differ in several key aspects from the cells of other eukaryotic organisms. Their distinctive features include:
- A large central vacuole, a water-filled volume enclosed by a membrane known as the tonoplast[1] that maintains the cell's turgor, controls movement of molecules between the cytosol and sap, stores useful material and digests waste proteins and organelles.
- A cell wall composed of cellulose and hemicellulose, pectin and in many cases lignin, is secreted by the protoplast on the outside of the cell membrane. This contrasts with the cell walls of fungi, which are made of chitin, and of bacteria, which are made of peptidoglycan. Cell walls perform many essential functions: they provide shape to form the tissue and organs of the plant, and play an important role in intercellular communication and plant-microbe interactions[2]
- Specialized cell-to-cell communication pathways known as plasmodesmata,[3] pores in the primary cell wall through which the plasmalemma and endoplasmic reticulum[4] of adjacent cells are continuous.
- Plastids, the most notable being the chloroplast, which contains chlorophyll, a green-colored pigment that absorbs sunlight, and allows the plant to make its own food in the process known as photosynthesis. Other types of plastids are the amyloplasts, specialized for starch storage, elaioplasts specialized for fat storage, and chromoplasts specialized for synthesis and storage of pigments. As in mitochondria, which have a genome encoding 37 genes,[5] plastids have their own genomes of about 100–120 unique genes[6] and, it is presumed, arose as prokaryotic endosymbionts living in the cells of an early eukaryotic ancestor of the land plants and algae.[7]
- Cell division by construction of a phragmoplast as a template for building a cell plate late in cytokinesis is characteristic of land plants and a few groups of algae, notably the Charophytes[8] and the Order Trentepohliales[9]
- The sperm of bryophytes and pteridophytes, cycads and Ginkgo have flagella[10] similar to those in animal cells,[11][12] but conifers and flowering plants lack flagella and centrioles.[13]
Cell types
- Parenchyma cells are living cells that have functions ranging from storage and support to photosynthesis and phloem loading (transfer cells). Apart from the xylem and phloem in their vascular bundles, leaves are composed mainly of parenchyma cells. Some parenchyma cells, as in the epidermis, are specialized for light penetration and focusing or regulation of gas exchange, but others are among the least specialized cells in plant tissue, and may remain totipotent, capable of dividing to produce new populations of undifferentiated cells, throughout their lives. Parenchyma cells have thin, permeable primary walls enabling the transport of small molecules between them, and their cytoplasm is responsible for a wide range of biochemical functions such as nectar secretion, or the manufacture of secondary products that discourage herbivory. Parenchyma cells that contain many chloroplasts and are concerned primarily with photosynthesis are called chlorenchyma cells. Others, such as the majority of the parenchyma cells in potato tubers and the seed cotyledons of legumes, have a storage function.
- Collenchyma cells – collenchyma cells are alive at maturity and have only a primary wall. These cells mature from meristem derivatives that initially resemble parenchyma, but differences quickly become apparent. Plastids do not develop, and the secretory apparatus (ER and Golgi) proliferates to secrete additional primary wall. The wall is most commonly thickest at the corners, where three or more cells come in contact, and thinnest where only two cells come in contact, though other arrangements of the wall thickening are possible.[14]
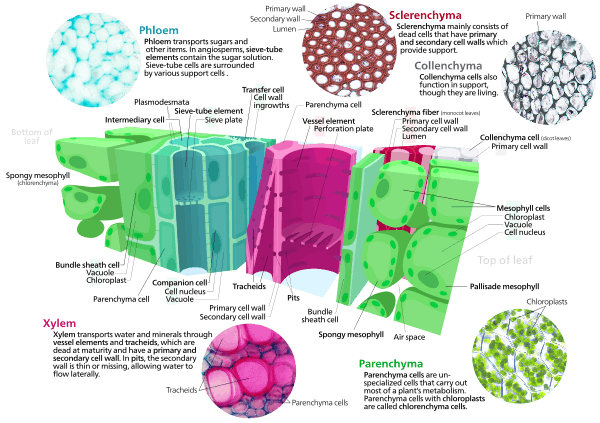
Pectin and hemicellulose are the dominant constituents of collenchyma cell walls of dicotyledon angiosperms, which may contain as little as 20% of cellulose in Petasites.[15] Collenchyma cells are typically quite elongated, and may divide transversely to give a septate appearance. The role of this cell type is to support the plant in axes still growing in length, and to confer flexibility and tensile strength on tissues. The primary wall lacks lignin that would make it tough and rigid, so this cell type provides what could be called plastic support – support that can hold a young stem or petiole into the air, but in cells that can be stretched as the cells around them elongate. Stretchable support (without elastic snap-back) is a good way to describe what collenchyma does. Parts of the strings in celery are collenchyma.
- Sclerenchyma cells – Sclerenchyma cells (from the Greek skleros, hard) are hard and tough cells with a function in mechanical support. They are of two broad types – sclereids or stone cells and fibres. The cells develop an extensive secondary cell wall that is laid down on the inside of the primary cell wall. The secondary wall is impregnated with lignin, making it hard and impermeable to water. Thus, these cells cannot survive for long' as they cannot exchange sufficient material to maintain active metabolism. Sclerenchyma cells are typically dead at functional maturity, and the cytoplasm is missing, leaving an empty central cavity.
Functions for sclereid cells (hard cells that give leaves or fruits a gritty texture) include discouraging herbivory, by damaging digestive passages in small insect larval stages, and physical protection (a solid tissue of hard sclereid cells form the pit wall in a peach and many other fruits). Functions of fibres include provision of load-bearing support and tensile strength to the leaves and stems of herbaceous plants.[14] Sclerenchyma fibres are not involved in conduction, either of water and nutrients (as in the xylem) or of carbon compounds (as in the phloem), but it is likely that they may have evolved as modifications of xylem and phloem initials in early land plants.
Tissue types

The major classes of cells differentiate from undifferentiated meristematic cells (analogous to the stem cells of animals) to form the tissue structures of roots, stems, leaves, flowers, and reproductive structures.
Xylem cells[16] are elongated cells with lignified secondary thickening of the cell walls. Xylem cells are specialised for conduction of water, and first appeared in plants during their transition to land in the Silurian period more than 425 million years ago (see Cooksonia). The possession of xylem defines the vascular plants or Tracheophytes. Xylem tracheids are pointed, elongated xylem cells, the simplest of which have continuous primary cell walls and lignified secondary wall thickenings in the form of rings, hoops, or reticulate networks. More complex tracheids with valve-like perforations called bordered pits characterise the gymnosperms. The ferns and other pteridophytes and the gymnosperms have only xylem tracheids, while the angiosperms also have xylem vessels. Vessel members are hollow xylem cells without end walls that are aligned end-to-end so as to form long continuous tubes. The bryophytes lack true xylem cells, but their sporophytes have a water-conducting tissue known as the hydrome that is composed of elongated cells of simpler construction.
Phloem is a specialised tissue for food transport in higher plants. Phloem cells mainly transport sucrose along pressure gradients generated by osmosis. This phenomenon is called translocation. Phloem consists of two cell types, the sieve tubes and the intimately associated companion cells. The sieve tube elements lack nuclei and ribosomes, and their metabolism and functions are regulated by the adjacent nucleate companion cells. Sieve tubes are joined end-to-end with perforate end-plates between known as sieve plates, which allow transport of photosynthate between the sieve elements. The companion cells, connected to the sieve tubes via plasmodesmata, are responsible for loading the phloem with sugars. The bryophytes lack phloem, but moss sporophytes have a simpler tissue with analogous function known as the leptome.

Plant epidermal cells are specialised parenchyma cells covering the external surfaces of leaves, stems and roots. The epidermal cells of aerial organs arise from the superficial layer of cells known as the tunica (L1 and L2 layers) that covers the plant shoot apex,[14] whereas the cortex and vascular tissues arise from innermost layer of the shoot apex known as the corpus (L3 layer). The epidermis of roots originates from the layer of cells immediately beneath the root cap.
The epidermis of all aerial organs, but not roots, is covered with a cuticle made of the polyester cutin and/or the hydrocarbon polymer cutan with a superficial layer of epicuticular waxes. The epidermal cells of the primary shoot are thought to be the only plant cells with the biochemical capacity to synthesize cutin.[17] Several cell types may be present in the epidermis. Notable among these are the stomatal guard cells, glandular and clothing hairs or trichomes, and the root hairs of primary roots. In the shoot epidermis of most plants, only the guard cells have chloroplasts. Chloroplasts contain the green pigment chlorophyll which is needed for photosynthesis.
Organelles
References
- ↑ Raven, JA (1997). "The vacuole: a cost-benefit analysis". Advances in Botanical Research. 25: 59–86. doi:10.1016/S0065-2296(08)60148-2.
- ↑ Keegstra, K (2010). "Plant cell walls". Plant Physiology. 154 (2): 483–486. doi:10.1104/pp.110.161240.
- ↑ Oparka, KJ (1993). "Signalling via plasmodesmata-the neglected pathway". Seminars in Cell Biology. 4: 131–138. doi:10.1006/scel.1993.1016.
- ↑ Hepler, PK (1982). "Endoplasmic reticulum in the formation of the cell plate and plasmodesmata". Protoplasma. 111: 121–133. doi:10.1007/BF01282070.
- ↑ Anderson, S; Bankier, AT; Barrell, BG; de Bruijn, MH; Coulson, AR; Drouin, J; Eperon, IC; Nierlich, DP; Roe, BA; Sanger, F; Schreier, PH; Smith, AJ; Staden, R; Young, IG (1981). "Sequence and organization of the human mitochondrial genome". Nature. 290: 4–65. doi:10.1038/290457a0.
- ↑ Cui, L; Veeraraghavan, N; Richter, A; Wall, K; Jansen, RK; Leebens-Mack, J; Makalowska, I; dePamphilis, CW (2006). "ChloroplastDB: the chloroplast genome database". Nucleic Acids Research. 34: D692–696. doi:10.1093/nar/gkj055.
- ↑ Margulis, L (1970). Origin of eukaryotic cells. New Haven: Yale University Press. ISBN 978-0300013535.
- ↑ Lewis, LA; McCourt, RM (2004). "Green algae and the origin of land plants". American Journal of Botany. 91: 1535–1556. doi:10.3732/ajb.91.10.1535.
- ↑ López-Bautista, JM; Waters, DA; Chapman, RL (2003). "Phragmoplastin, green algae and the evolution of cytokinesis". International Journal of Systematic and Evolutionary Microbiology. 53: 1715–1718. doi:10.1099/ijs.0.02561-0.
- ↑ Silflow, CD; Lefebvre, PA (2001). "Assembly and motility of eukaryotic cilia and flagella. Lessons from Chlamydomonas reinhardtii". Plant Physiology. 127: 1500–1507. doi:10.1104/pp.010807.
- ↑ Manton, I; Clarke, B (1952). "An electron microscope study of the spermatozoid of Sphagnum". Journal of Experimental Botany. 3: 265–275. doi:10.1093/jxb/3.3.265.
- ↑ Paolillo, Jr., DJ (1967). "On the structure of the axoneme in flagella of Polytrichum juniperinum". Transactions of the American Microscopical Society. 86: 428–433.
- ↑ Raven, PH; Evert, RF; Eichhorm, SE (1999). Biology of Plants (6th ed.). New York: W.H. Freeman. ISBN 9780716762843.
- 1 2 3 Cutter, EG (1977). Plant Anatomy Part 1. Cells and Tissues. London: Edward Arnold. ISBN 0713126388.
- ↑ Roelofsen, PA (1959). The plant cell wall. Berlin: Gebrüder Borntraeger. ASIN B0007J57W0.
- ↑ MT Tyree; MH Zimmermann (2003) Xylem structure and the ascent of sap, 2nd edition, Springer-Verlag, New York USA
- ↑ Kolattukudy, PE (1996) Biosynthetic pathways of cutin and waxes, and their sensitivity to environmental stresses. In: Plant Cuticles. Ed. by G. Kerstiens, BIOS Scientific publishers Ltd., Oxford, pp 83–108
| Subdisciplines | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant groups | |||||||||||||||||||
| |||||||||||||||||||
| |||||||||||||||||||
| Plant growth and habit | |||||||||||||||||||
| Reproduction | |||||||||||||||||||
| Plant taxonomy | |||||||||||||||||||
| Practice | |||||||||||||||||||
| |||||||||||||||||||
| |||||||||||||||||||