CREB

CREB (cAMP response element-binding protein)[1] is a cellular transcription factor. It binds to certain DNA sequences called cAMP response elements (CRE), thereby increasing or decreasing the transcription of the downstream genes.[2] CREB was first described in 1987 as a cAMP-responsive transcription factor regulating the somatostatin gene.[3]
Genes whose transcription is regulated by CREB include: c-fos, BDNF, tyrosine hydroxylase, numerous neuropeptides (such as somatostatin, enkephalin, VGF, corticotropin-releasing hormone),[2] and genes involved in the mammalian circadian clock (PER1, PER2).[4]
CREB is closely related in structure and function to CREM (cAMP response element modulator) and ATF-1 (activating transcription factor-1) proteins. CREB proteins are expressed in many animals, including humans.
CREB has a well-documented role in neuronal plasticity and long-term memory formation in the brain and has been shown to be integral in the formation of spatial memory.[5] CREB downregulation is implicated in the pathology of Alzheimer's disease and increasing the expression of CREB is being considered as a possible therapeutic target for Alzheimers disease.[6] CREB also has a role in photoentrainment in mammals.
Subtypes
The following genes encode CREB or CREB-like proteins:
- CREB1 (CREB1)
- CREB2 renamed ATF4 (ATF4)
- CREB3 (CREB3)
- CREB5 (CREB5)
- CREB3L1 (CREB3L1)
- CREB3L2 (CREB3L2)
- CREB3L3 (CREB3L3)
- CREB3L4 (CREB3L4)
Structure
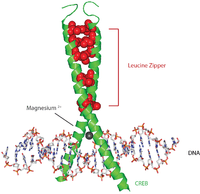
CREB proteins are activated by phosphorylation from various kinases, including PKA, and Ca2+/calmodulin-dependent protein kinases on the Serine 133 residue.[7] When activated, CREB protein recruits other transcriptional coactivators to bind to CRE promoter 5’ upstream region. Hydrophobic leucine amino acids are located along the inner edge of the alpha helix. These leucine residues tightly bind to leucine residues of another CREB protein forming the dimer. This chain of leucine residues forms the leucine zipper motif. The protein also has a magnesium ion that facilitates binding to DNA.
cAMP response element
The cAMP response element (CRE) is the response element for CREB which contains the highly conserved nucleotide sequence, 5'-TGACGTCA-3’. CRE sites are typically found upstream of genes, within the promoter or enhancer regions.[8] There are approximately 750,000 palindromic and half-site CREs in the human genome. However, the majority of these sites remain unbound due to cytosine methylation[9] which physically obstructs protein binding.
Mechanism of action
A typical (albeit somewhat simplified) sequence of events is as follows: A signal arrives at the cell surface, activates the corresponding receptor, which leads to the production of a second messenger such as cAMP or Ca2+, which in turn activates a protein kinase. This protein kinase translocates to the cell nucleus, where it activates a CREB protein. The activated CREB protein then binds to a CRE region, and is then bound to by CBP (CREB-binding protein), which coactivates it, allowing it to switch certain genes on or off. The DNA binding of CREB is mediated via its basic leucine zipper domain (bZIP domain) as depicted in the image.
Function
CREB has many functions in many different organs, and some of its functions have been studied in relation to the brain.[10] CREB proteins in neurons are thought to be involved in the formation of long-term memories; this has been shown in the marine snail Aplysia, the fruit fly Drosophila melanogaster, in rats and in mice (see CREB in Molecular and Cellular Cognition).[1] CREB is necessary for the late stage of long-term potentiation. CREB also has an important role in the development of drug addiction and even more so in psychological dependence.[11][12][13] There are activator and repressor forms of CREB. Flies genetically engineered to overexpress the inactive form of CREB lose their ability to retain long-term memory. CREB is also important for the survival of neurons, as shown in genetically engineered mice, where CREB and CREM were deleted in the brain. If CREB is lost in the whole developing mouse embryo, the mice die immediately after birth, again highlighting the critical role of CREB in promoting survival.
Disease linkage
Disturbance of CREB function in brain can contribute to the development and progression of Huntington's Disease.
Abnormalities of a protein that interacts with the KID domain of CREB, the CREB-binding protein, (CBP) is associated with Rubinstein-Taybi syndrome.
There is some evidence to suggest that the under-functioning of CREB is associated with Major Depressive Disorder.[14] Depressed rats with an overexpression of CREB in the dentate gyrus behaved similarly to rats treated with antidepressants.[15] From post-mortem examinations it has also been shown that the cortices of patients with untreated major depressive disorder contain reduced concentrations of CREB compared to both healthy controls and patients treated with antidepressants.[15] The function of CREB can be modulated via a signalling pathway resulting from the binding of serotonin and noradrenaline to post-synaptic G-protein coupled receptors. Dysfunction of these neurotransmitters is also implicated in major depressive disorder.[14]
CREB is also thought to be involved in the growth of some types of cancer.
Involvement in Circadian Rhythms
Entrainment of the mammalian circadian clock is established via light induction of PER. Light excites melanopsin-containing photosensitive retinal ganglion cells which signal to the suprachiasmatic nucleus (SCN) via the Retinohypothalamic tract (RHT). Excitation of the RHT signals the release of glutamate which is received by NMDA receptors on SCN, resulting in a calcium influx into the SCN. Calcium induces the activity of Ca2+/calmodulin-dependent protein kinases, resulting in the activation of PKA, PKC, and CK2.[16] These kinases then phosphorylate CREB in a circadian manner that further regulates downstream gene expression.[17] The phosphorylated CREB recognizes the cAMP Response Element and serves as a transcription factor for Per1 and Per2, two genes that regulate the mammalian circadian clock. This induction of PER protein can entrain the circadian clock to light/dark cycles inhibits its own transcription via a transcription-translation feedback loop which can advance or delay the circadian clock. However, the responsiveness of PER1 and PER2 protein induction is only significant during the subjective night.[4]
Discovery of CREB involvement in circadian rhythms
Michael Greenberg first demonstrated the role of CREB in the mammalian circadian clock in 1993 through a series of experiments that correlated phase-specific light pulses with CREB phosphorylation. In vitro, light during the subjective night increased phosphorylation of CREB rather than CREB protein levels. In vivo, phase shift-inducing light pulses during the subjective night correlated with CREB phosphorylation in the SCN.[18] Experiments by Gunther Schutz in 2002 demonstrated that mutant mice lacking the Ser142 phosphorylation site failed to induce the clock regulatory gene mPer1 in response to a light pulse. Furthermore, these mutant mice had difficulty entraining to light-dark cycles.[19]
See also
References
- 1 2 Bourtchuladze et al. "Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein." Cell 79(1):59-68. PMID 7923378
- 1 2 Purves, Dale; George J. Augustine; David Fitzpatrick; William C. Hall; Anthony-Samuel LaMantia; James O. McNamara & Leonard E. White (2008). Neuroscience (4th ed.). Sinauer Associates. pp. 170–6. ISBN 978-0-87893-697-7.
- ↑ Binding of a nuclear protein to the cyclic-AMP response element of the somatostatin gene. Montminy MR and Bilezikjian LM. Nature. 1987 Jul 9-15;328(6126):175-8.
- 1 2 Dibner, Charna; Schibler, Ueli; Albrecht, Urs (2010). "The Mammalian Circadian Timing System: Organization and Coordination of Central and Peripheral Clocks". Annual Review of Physiology. 72 (1): 517–549. doi:10.1146/annurev-physiol-021909-135821. PMID 20148687. Retrieved 2015-04-09.
- ↑ Silva; et al. "CREB and Memory" (PDF). Annual Review of Neuroscience. 21: 127–148. doi:10.1146/annurev.neuro.21.1.127.
- ↑ Downregulation of CREB expression in Alzheimer’s brain and in Ab-treated rat hippocampal neurons
- ↑ Shaywitz, Adam J.; Greenberg, Michael E. (1999). "CREB: A Stimulus-Induced Transcription Factor Activated by A Diverse Array of Extracellular Signals". Annual Review of Biochemistry. 68 (1): 821–861. doi:10.1146/annurev.biochem.68.1.821. PMID 10872467. Retrieved 2015-04-09.
- ↑ "The many faces of CREB". linkinghub.elsevier.com. Retrieved 2015-04-09.
- ↑ Altarejos, Judith Y.; Montminy, Marc (March 2011). "CREB and the CRTC co-activators: sensors for hormonal and metabolic signals". Nature Reviews Molecular Cell Biology. 12 (3): 141–151. doi:10.1038/nrm3072. ISSN 1471-0072. PMC 4324555
 . PMID 21346730. Retrieved 2015-04-09.
. PMID 21346730. Retrieved 2015-04-09. - ↑ Carlezon WA, Duman RS, Nestler EJ (August 2005). "The many faces of CREB". Trends in Neurosciences. 28 (8): 436–45. doi:10.1016/j.tins.2005.06.005. PMID 15982754.
- ↑ Nazarian A, Sun WL, Zhou L, Kemen LM, Jenab S, Quinones-Jenab V (April 2009). "Sex differences in basal and cocaine-induced alterations in PKA and CREB proteins in the nucleus accumbens". Psychopharmacology. 203 (3): 641–50. doi:10.1007/s00213-008-1411-5. PMID 19052730.
- ↑ Wang Y, Ghezzi A, Yin JC, Atkinson NS (June 2009). "CREB regulation of BK channel gene expression underlies rapid drug tolerance". Genes, Brain, and Behavior. 8 (4): 369–76. doi:10.1111/j.1601-183X.2009.00479.x. PMC 2796570. PMID 19243452.
- ↑ DiRocco DP, Scheiner ZS, Sindreu CB, Chan GC, Storm DR (February 2009). "A role for calmodulin-stimulated adenylyl cyclases in cocaine sensitization". Journal of Neuroscience. 29 (8): 2393–403. doi:10.1523/JNEUROSCI.4356-08.2009. PMC 2678191. PMID 19244515.
- 1 2 Belmaker, R. H., and Galila Agam. "Major depressive disorder." New England Journal of Medicine 358.1 (2008): 55-68.
- 1 2 Blendy JA. The role of CREB in depression and antidepressant treatment. Biol Psychiatry 2006;59:1144-50.
- ↑ Iyer, Rajashekar; Wang, Tongfei; Gillette, Martha (September 19, 2014). "Circadian gating of neuronal functionality: a basis for iterative metaplasticity". Frontiers in Systems Neuroscience. 8: 164. doi:10.3389/fnsys.2014.00164. PMC 4168688. PMID 25285070.
- ↑ Obrietan, Karl; Impey, Soren; Smith, Dave; Athos, Jaime; Storm, Derrick R. (April 11, 2002). "Circadian regulation of cAMP response element-mediated gene expression in the suprachiasmatic nuclei.". Neuron. 274: 17748–17756. doi:10.1074/jbc.274.25.17748.
- ↑ Ginty, D. D.; Kornhauser, J. M.; Thompson, M. A.; Bading, H.; Mayo, K. E.; Takahashi, J. S.; Greenberg, M. E. (April 9, 1993). "Regulation of CREB phosphorylation in the suprachiasmatic nucleus by light and a circadian clock". Science. 260 (5105): 238–241. doi:10.1126/science.8097062. ISSN 0036-8075. PMID 8097062. Retrieved April 8, 2015.
- ↑ Gau, Daniel; Lemberger, Thomas; von Gall, Charlotte; Kretz, Oliver; Le Minh, Nguyet; Gass, Peter; Schmid, Wolfgang; Schibler, Ueli; Korf, Horst W. (April 11, 2002). "Phosphorylation of CREB Ser142 Regulates Light-Induced Phase Shifts of the Circadian Clock". Neuron. 34: 245–253. doi:10.1016/S0896-6273(02)00656-6.
- Bibliography
- Lauren Slater (2005). Opening Skinner's Box: Great Psychological Experiments of the Twentieth Century. New York: W. W. Norton & Company. ISBN 0-393-32655-1.
- Barco A, Bailey C, Kandel E (2006). "Common molecular mechanisms in explicit and implicit memory". J. Neurochem. 97 (6): 1520–33. doi:10.1111/j.1471-4159.2006.03870.x. PMID 16805766.
- Conkright M, Montminy M (2005). "CREB: the unindicted cancer co-conspirator". Trends Cell Biol. 15 (9): 457–9. doi:10.1016/j.tcb.2005.07.007. PMID 16084096.
- Mantamadiotis T, Lemberger T, Bleckmann S, Kern H, Kretz O, Martin Villalba A, Tronche F, Kellendonk C, Gau D, Kapfhammer J, Otto C, Schmid W, Schütz G (2002). "Disruption of CREB function in brain leads to neurodegeneration". Nat. Genet. 31 (1): 47–54. doi:10.1038/ng882. PMID 11967539.
- Mayr B, Montminy M (2001). "Transcriptional regulation by the phosphorylation-dependent factor CREB". Nat. Rev. Mol. Cell Biol. 2 (8): 599–609. doi:10.1038/35085068. PMID 11483993.
- Yin J, Del Vecchio M, Zhou H, Tully T (1995). "CREB as a memory modulator: induced expression of a dCREB2 activator isoform enhances long-term memory in Drosophila". Cell. 81 (1): 107–15. doi:10.1016/0092-8674(95)90375-5. PMID 7720066.
- Yin J, Wallach J, Del Vecchio M, Wilder E, Zhou H, Quinn W, Tully T (1994). "Induction of a dominant negative CREB transgene specifically blocks long-term memory in Drosophila". Cell. 79 (1): 49–58. doi:10.1016/0092-8674(94)90399-9. PMID 7923376.
External links
- http://www.ebi.ac.uk/interpro/entry/IPR001630
- Johannessen, M., Pedersen Delghandi, M., and Moens, U. (2004) - What Turns CREB on ? - Cell Signall.; 10:1211-1227. http://www.sigtrans.org/publications/what-turns-creb-on/
- http://focus.hms.harvard.edu/2001/Oct26_2001/neuroscience.html
- CREB Protein at the US National Library of Medicine Medical Subject Headings (MeSH)
- Drosophila Cyclic-AMP response element binding protein A - The Interactive Fly
- Drosophila Cyclic-AMP response element binding protein B at 17A - The Interactive Fly